Palomares-Rius Juan E, Escobar Carolina, Cabrera Javier, Vovlas Alessio, Castillo Pablo
Department of Crop Protection, Institute for Sustainable Agriculture (CSIC), Córdoba, Spain.
Plant Biotechnology and Molecular Biology Group, University of Castilla La Mancha, Toledo, Spain.
Front Plant Sci. 2017 Nov 16;8:1987. doi: 10.3389/fpls.2017.01987. eCollection 2017.
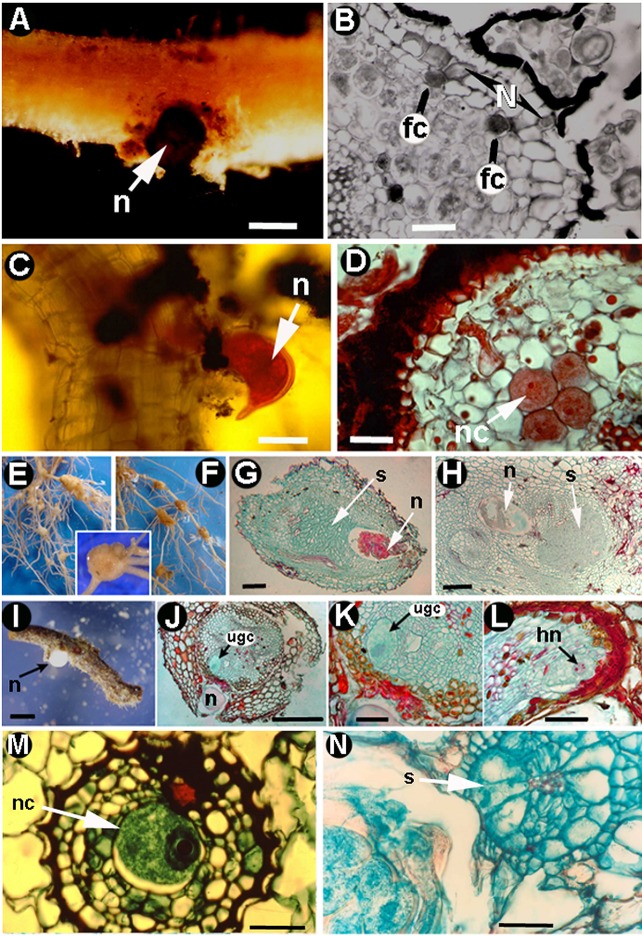
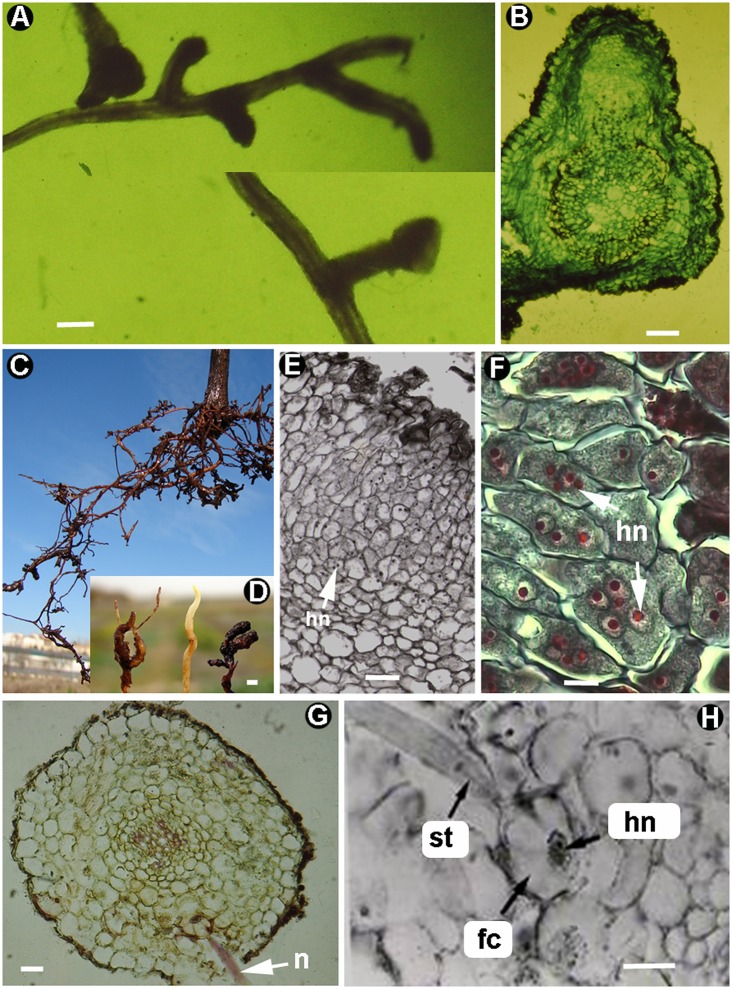
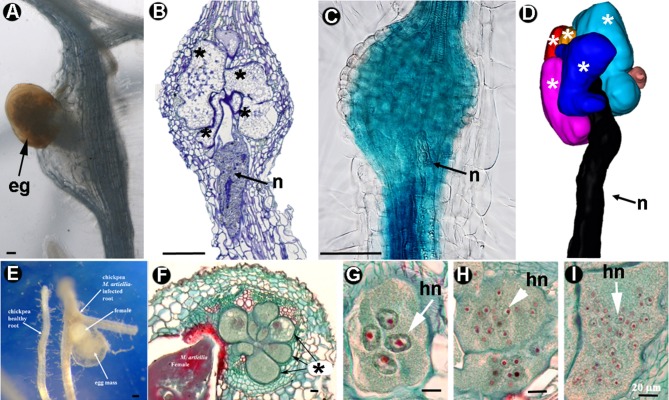
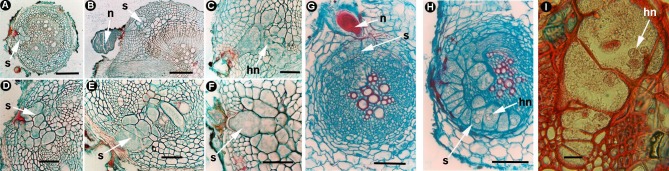
Plant-parasitic nematodes (PPNs) interact with plants in different ways, for example, through subtle feeding behavior, migrating destructively through infected tissues, or acting as virus-vectors for nepoviruses. They are all obligate biotrophic parasites as they derive their nutrients from living cells which they modify using pharyngeal gland secretions prior to food ingestion. Some of them can also shield themselves against plant defenses to sustain a relatively long lasting interaction while feeding. This paper is centered on cell types or organs that are newly induced in plants during PPN parasitism, including recent approaches to their study based on molecular biology combined with cell biology-histopathology. This issue has already been reviewed extensively for major PPNs (i.e., root-knot or cyst nematodes), but not for other genera (viz. spp.). PPNs have evolved with plants and this co-evolution process has allowed the induction of new types of plant cells necessary for their parasitism. There are four basic types of feeding cells: (i) non-hypertrophied nurse cells; (ii) single giant cells; (iii) syncytia; and (iv) coenocytes. Variations in the structure of these cells within each group are also present between some genera depending on the nematode species viz. or . This variability of feeding sites may be related in some way to PPN life style (migratory ectoparasites, sedentary ectoparasites, migratory ecto-endoparasites, migratory endoparasites, or sedentary endoparasites). Apart from their co-evolution with plants, the response of plant cells and roots are closely related to feeding behavior, the anatomy of the nematode (mainly stylet size, which could reach different types of cells in the plant), and the secretory fluids produced in the pharyngeal glands. These secretory fluids are injected through the stylet into perforated cells where they modify plant cytoplasm prior to food removal. Some species do not produce specialized feeding sites (viz. ), but may develop a specialized modification of the root system (e.g., unspecialized root galls or a profusion of roots). This review introduces new data on cell types and plant organs stimulated by PPNs using sources varying from traditional histopathology to new holistic methodologies.
植物寄生线虫(PPNs)以不同方式与植物相互作用,例如,通过微妙的取食行为、在受感染组织中破坏性迁移,或作为线虫传多面体病毒的病毒载体。它们都是专性活体营养型寄生虫,因为它们从活细胞中获取营养,并在摄取食物之前利用咽腺分泌物对活细胞进行改造。其中一些线虫还能抵御植物防御,以便在取食时维持相对持久的相互作用。本文聚焦于PPNs寄生期间植物新诱导产生的细胞类型或器官,包括基于分子生物学结合细胞生物学 - 组织病理学的最新研究方法。关于主要的PPNs(即根结线虫或孢囊线虫),这个问题已经有了广泛的综述,但对于其他属(即 spp.)尚未有相关综述。PPNs与植物共同进化,这种共同进化过程使得诱导出其寄生所需的新型植物细胞成为可能。有四种基本类型的取食细胞:(i)非肥大性滋养细胞;(ii)单个巨型细胞;(iii)多核细胞;(iv)多核体。根据线虫种类(即 或 ),每组内这些细胞的结构在某些属之间也存在差异。取食部位的这种变异性可能在某种程度上与PPN的生活方式(迁移性外寄生线虫、固定性外寄生线虫、迁移性内外寄生线虫、迁移性内寄生线虫或固定性内寄生线虫)有关。除了与植物共同进化外,植物细胞和根的反应与取食行为、线虫的解剖结构(主要是口针大小,它可以到达植物中的不同类型细胞)以及咽腺产生的分泌液密切相关。这些分泌液通过口针注入穿孔细胞,在去除食物之前改变植物细胞质。一些物种不会产生特化的取食部位(即 ),但可能会使根系发生特化改变(例如,非特化的根瘤或大量的根)。本综述利用从传统组织病理学到新的整体方法等不同来源的数据,介绍了关于PPNs刺激的细胞类型和植物器官的新数据。