Department of Chemical and Systems Biology, Stanford School of Medicine, Stanford University, Stanford, California 94305, USA.
Department of Genetics, Stanford School of Medicine, Stanford University, Stanford, California 94305, USA.
Nature. 2018 Jan 11;553(7687):228-232. doi: 10.1038/nature25179. Epub 2017 Dec 6.
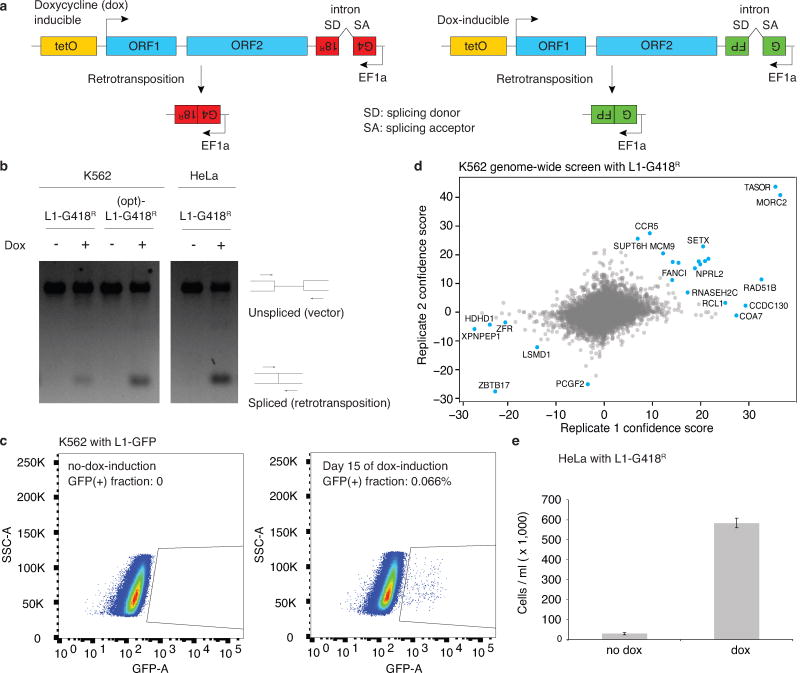
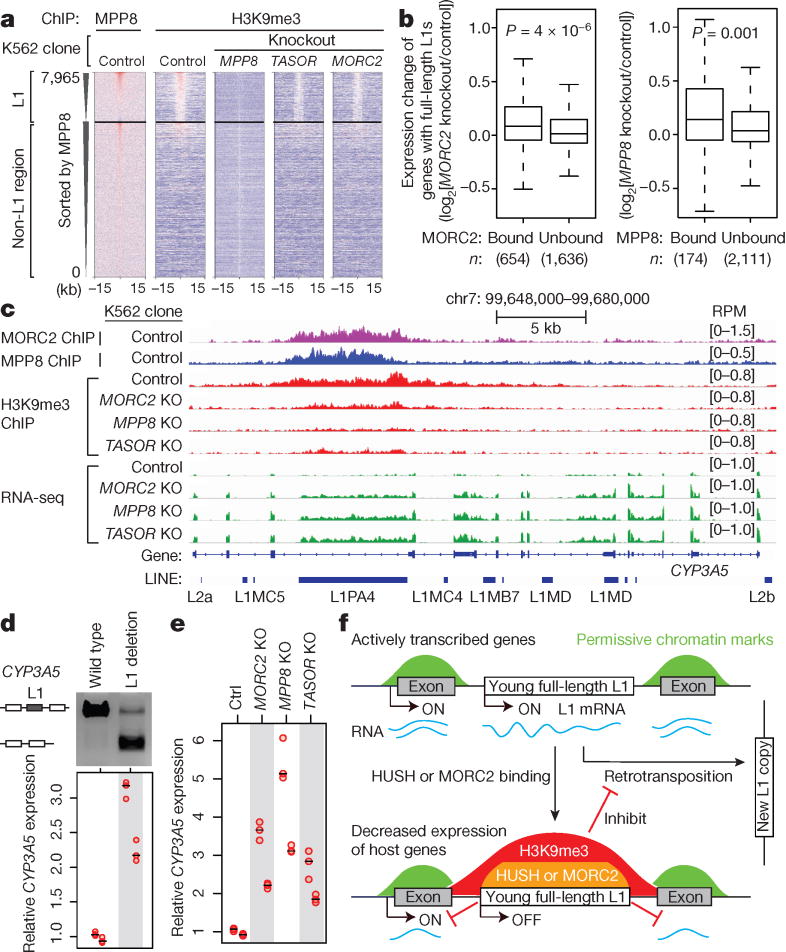
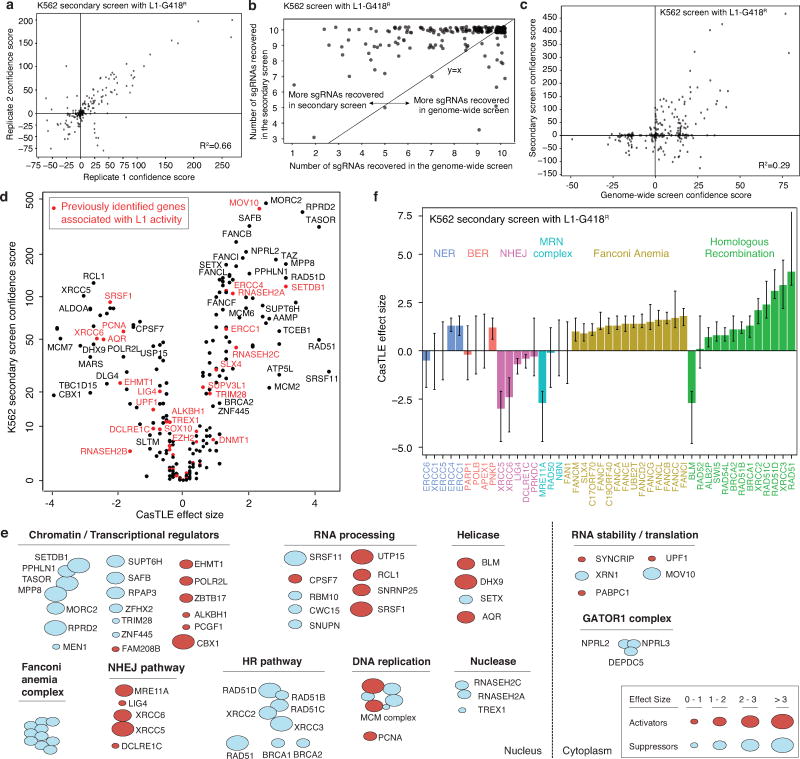
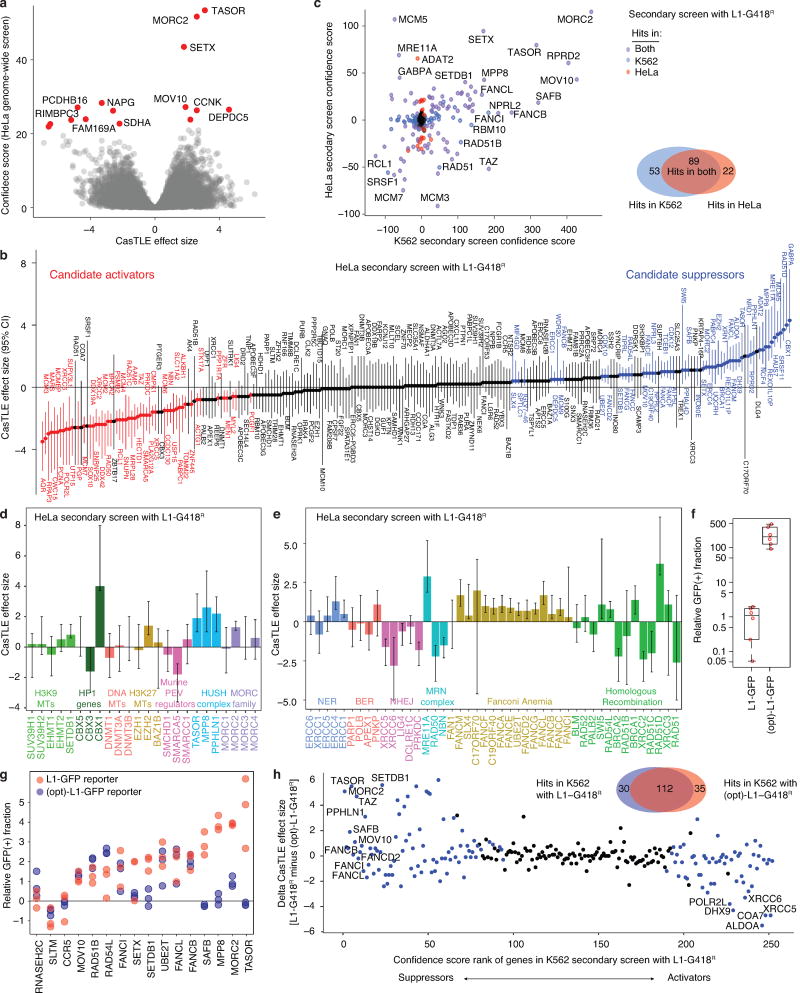
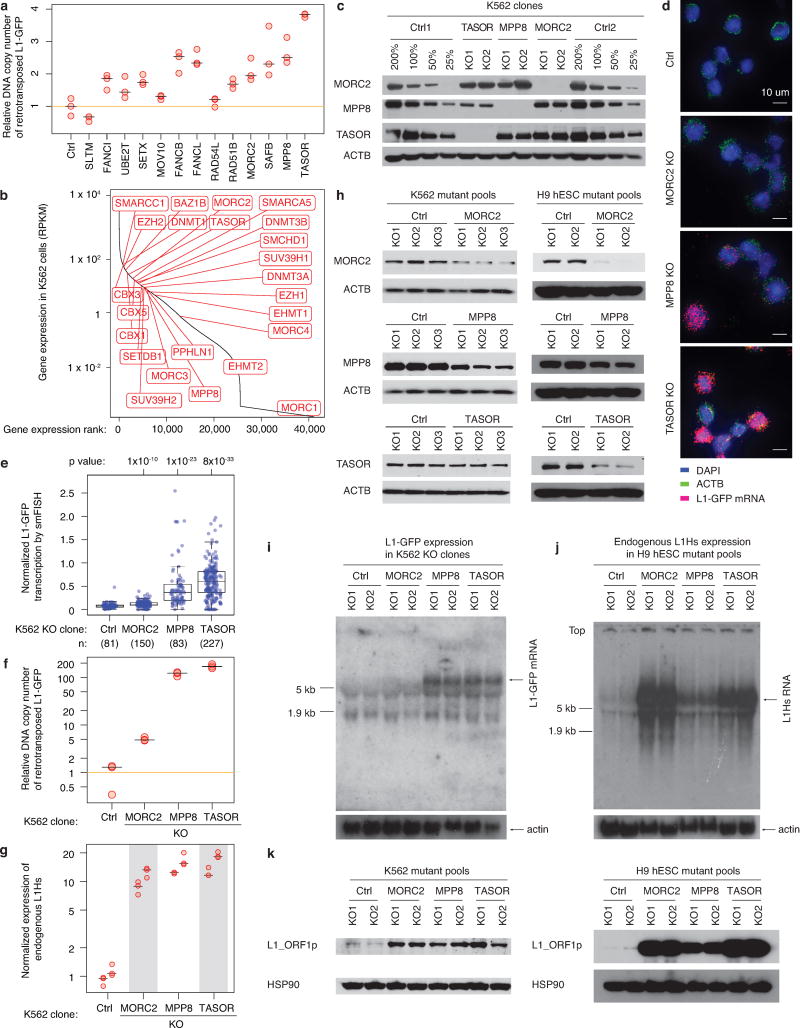
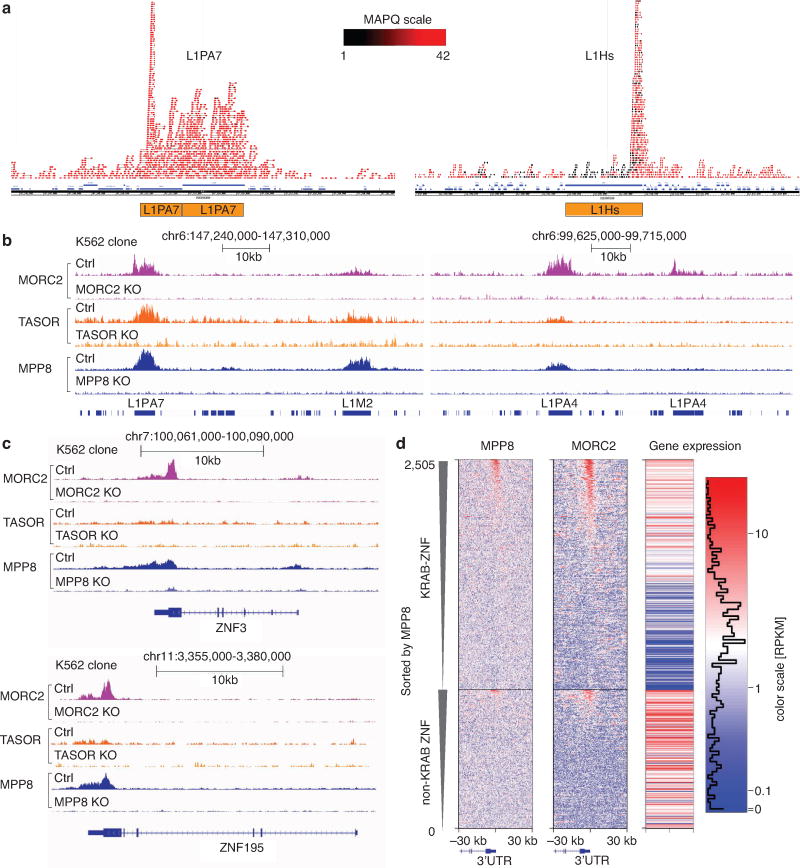
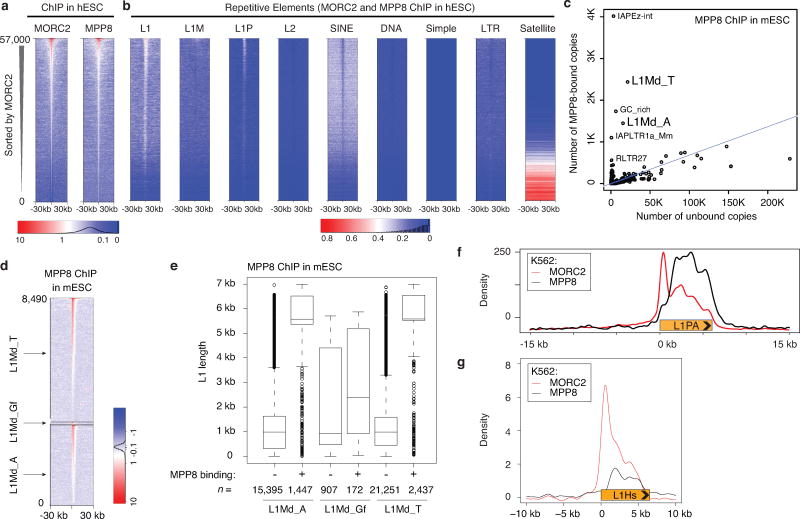
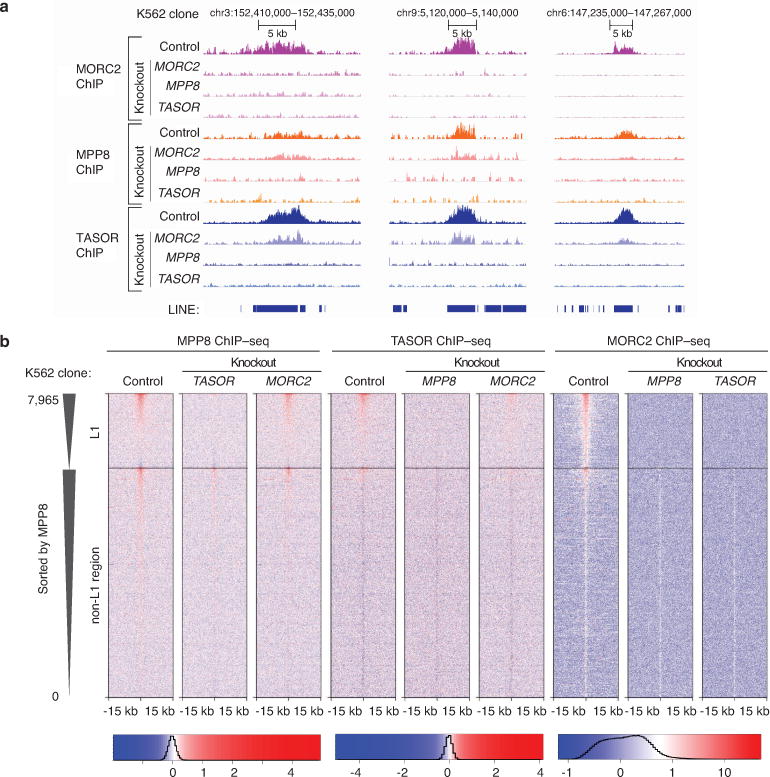
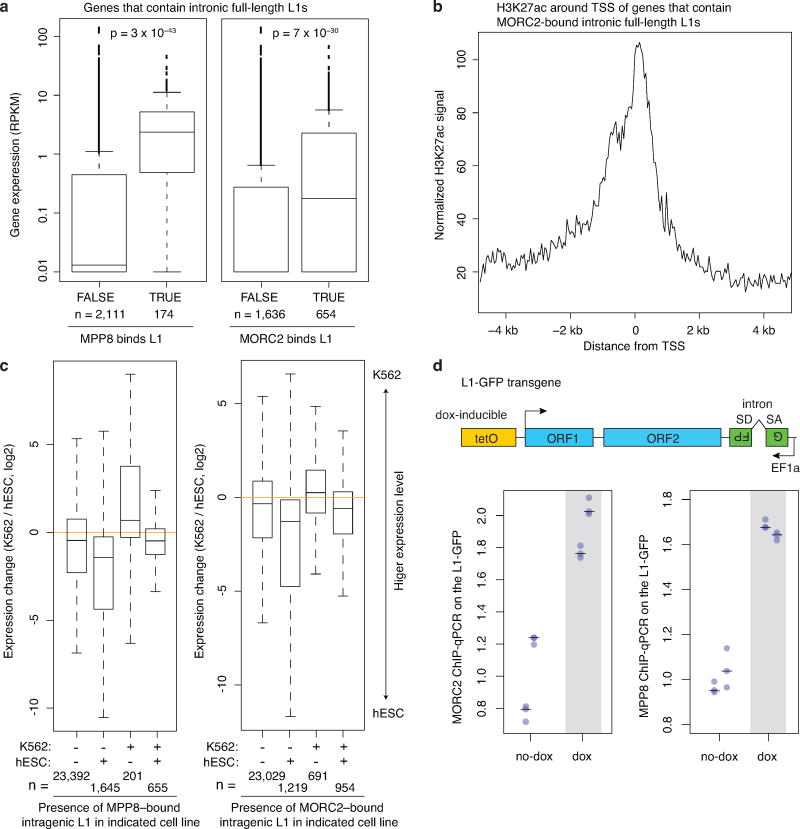
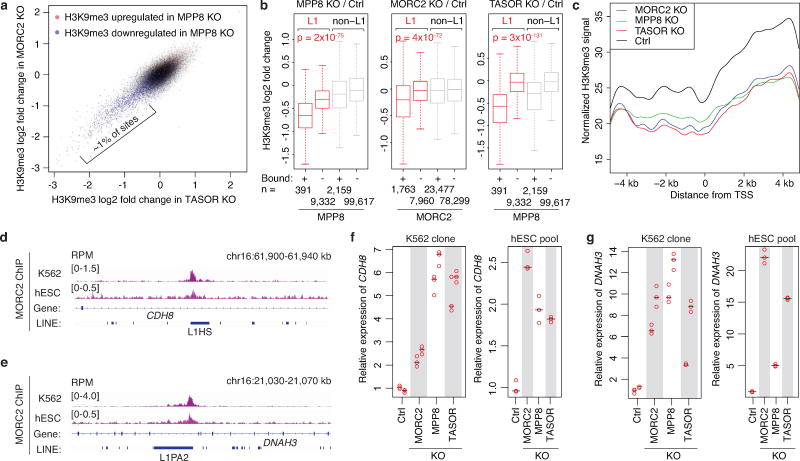
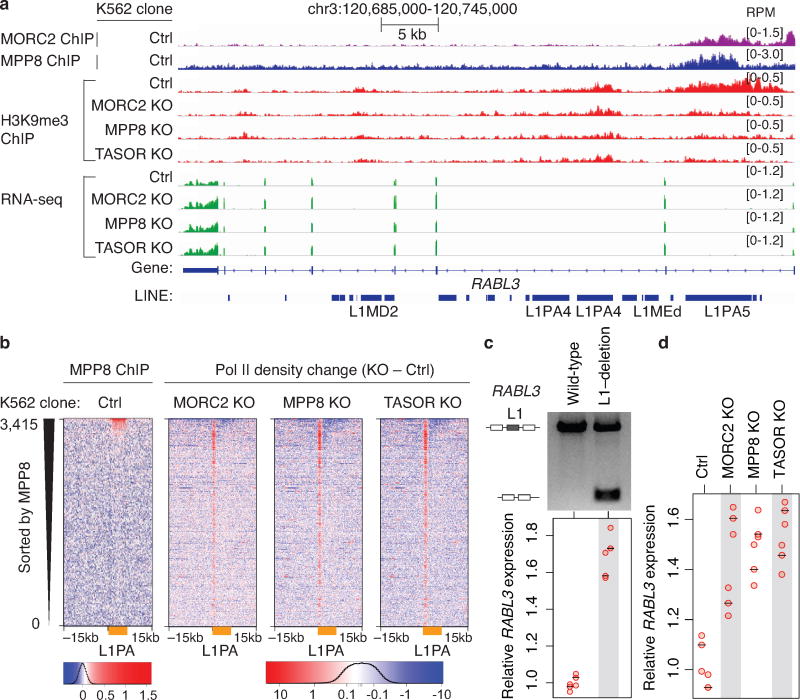
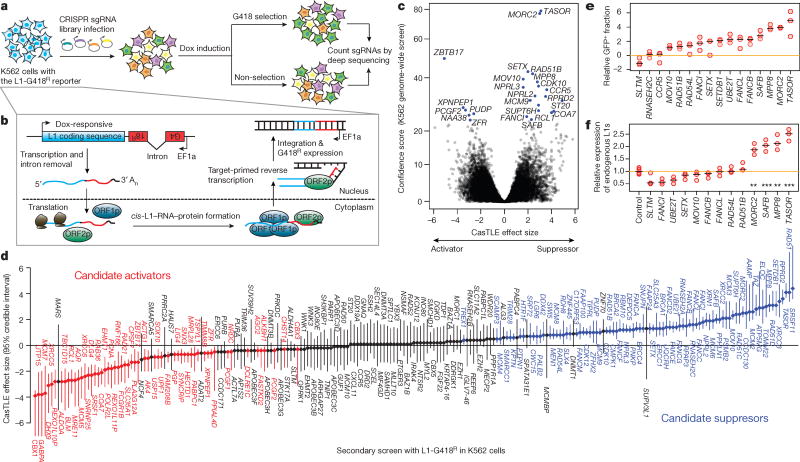
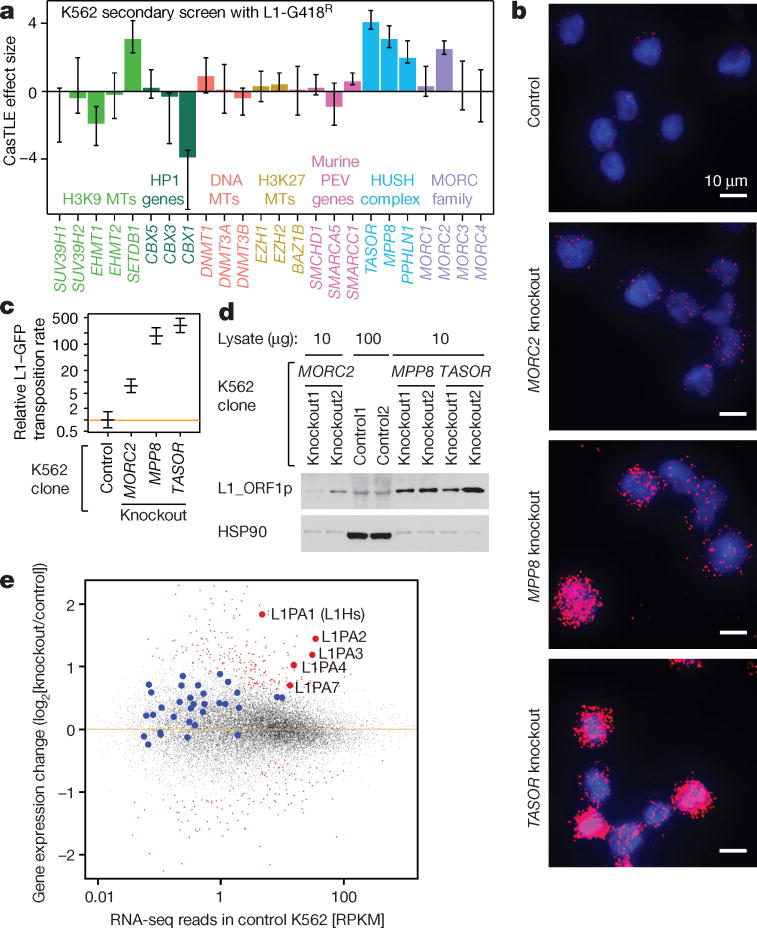
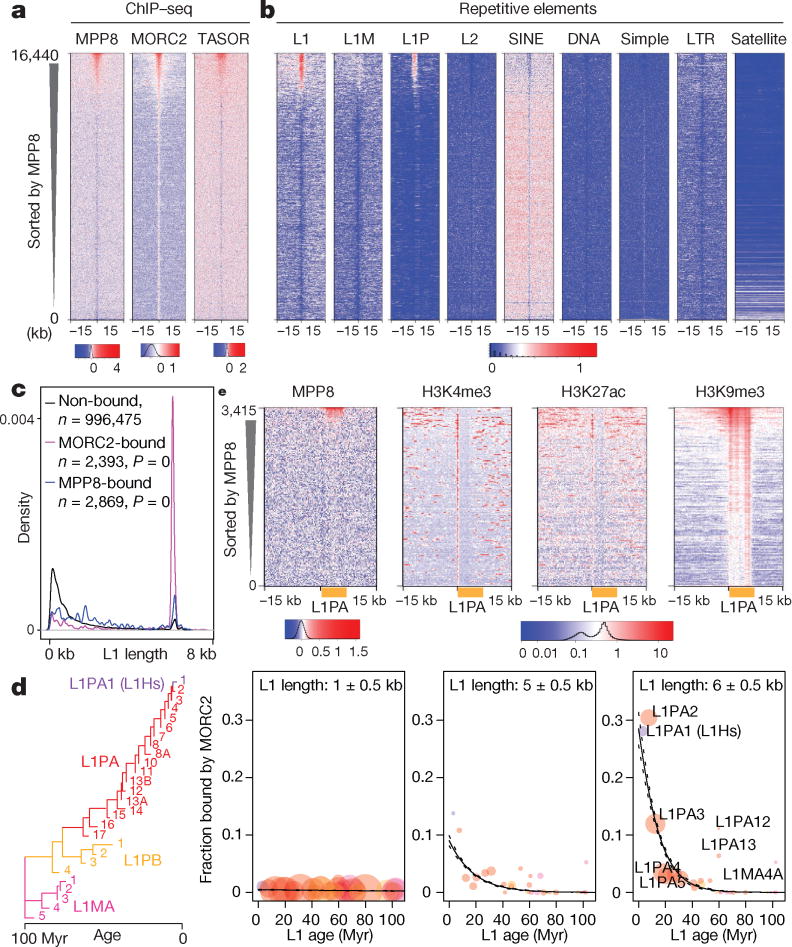
Transposable elements, also known as transposons, are now recognized not only as parasitic DNA, the spread of which in the genome must be controlled by the host, but also as major players in genome evolution and regulation. Long interspersed element-1 (LINE-1, also known as L1), the only currently autonomous mobile transposon in humans, occupies 17% of the genome and generates inter- and intra-individual genetic variation, in some cases resulting in disease. However, how L1 activity is controlled and the function of L1s in host gene regulation are not completely understood. Here we use CRISPR-Cas9 screening strategies in two distinct human cell lines to provide a genome-wide survey of genes involved in the control of L1 retrotransposition. We identify functionally diverse genes that either promote or restrict L1 retrotransposition. These genes, which are often associated with human diseases, control the L1 life cycle at the transcriptional or the post-transcriptional level in a manner that can depend on the endogenous L1 nucleotide sequence, underscoring the complexity of L1 regulation. We further investigate the restriction of L1 by the protein MORC2 and by the human silencing hub (HUSH) complex subunits MPP8 and TASOR. HUSH and MORC2 can selectively bind evolutionarily young, full-length L1s located within transcriptionally permissive euchromatic environments, and promote deposition of histone H3 Lys9 trimethylation (H3K9me3) for transcriptional silencing. Notably, these silencing events often occur within introns of transcriptionally active genes, and lead to the downregulation of host gene expression in a HUSH-, MORC2-, and L1-dependent manner. Together, these results provide a rich resource for studies of L1 retrotransposition, elucidate a novel L1 restriction pathway and illustrate how epigenetic silencing of transposable elements rewires host gene expression programs.
转座元件,也被称为转座子,现在不仅被认为是寄生 DNA,其在基因组中的传播必须受到宿主的控制,而且还是基因组进化和调控的主要参与者。长散布元件-1(LINE-1,也称为 L1)是人类中唯一自主移动的转座子,占据基因组的 17%,并产生个体间和个体内的遗传变异,在某些情况下导致疾病。然而,L1 活性是如何被控制的,以及 L1 在宿主基因调控中的功能尚不完全清楚。在这里,我们使用两种不同的人类细胞系中的 CRISPR-Cas9 筛选策略,对参与控制 L1 反转录转座的基因进行了全基因组调查。我们鉴定了功能多样的基因,这些基因要么促进要么限制 L1 反转录转座。这些基因通常与人类疾病有关,以依赖于内源性 L1 核苷酸序列的方式在转录或转录后水平上控制 L1 的生命周期,突显出 L1 调控的复杂性。我们进一步研究了 MORC2 蛋白和 HUSH 复合物亚基 MPP8 和 TASOR 对 L1 的限制作用。HUSH 和 MORC2 可以选择性地结合位于转录允许的常染色质环境中的进化上年轻的全长 L1,并促进组蛋白 H3 赖氨酸 9 三甲基化(H3K9me3)的沉积,以进行转录沉默。值得注意的是,这些沉默事件通常发生在转录活跃基因的内含子中,并以 HUSH、MORC2 和 L1 依赖的方式导致宿主基因表达的下调。总之,这些结果为 L1 反转录转座的研究提供了丰富的资源,阐明了一种新的 L1 限制途径,并说明了转座元件的表观遗传沉默如何重编宿主基因表达程序。