Watson School of Biological Sciences, Cold Spring Harbor Laboratory, New York, United States.
Elife. 2017 Dec 20;6:e29005. doi: 10.7554/eLife.29005.
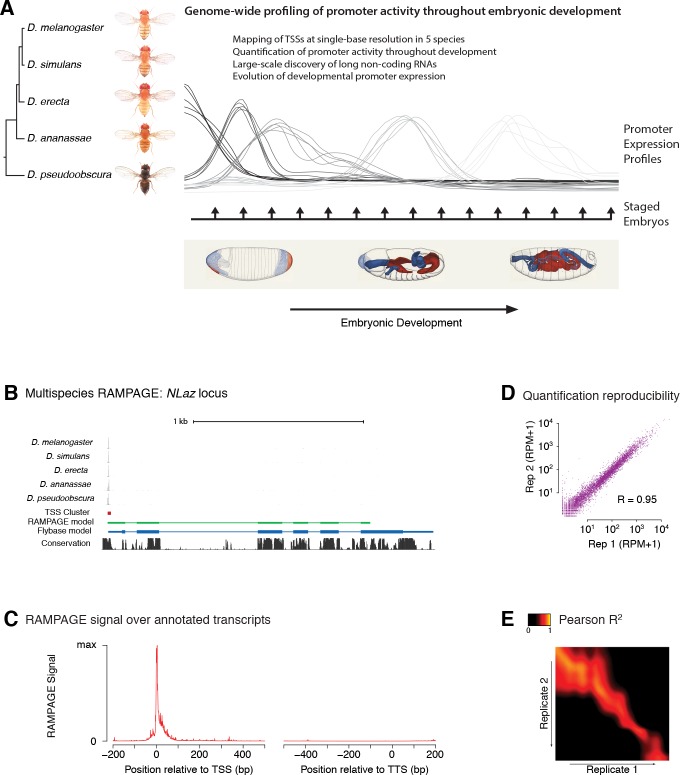
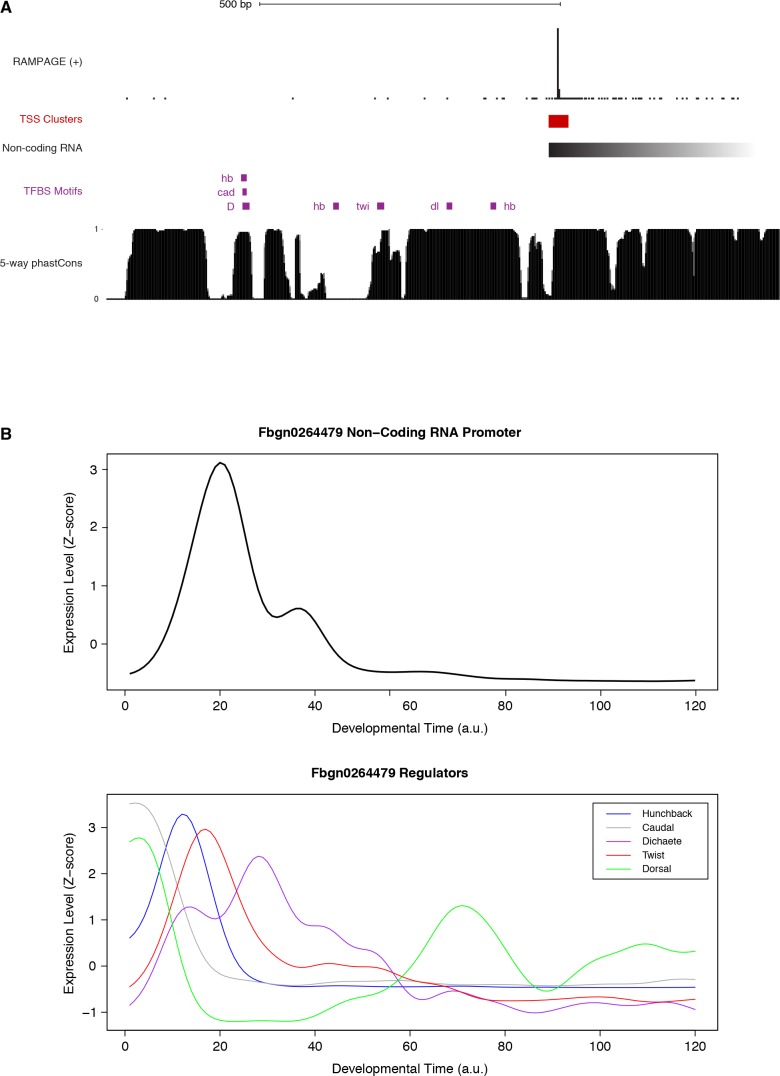
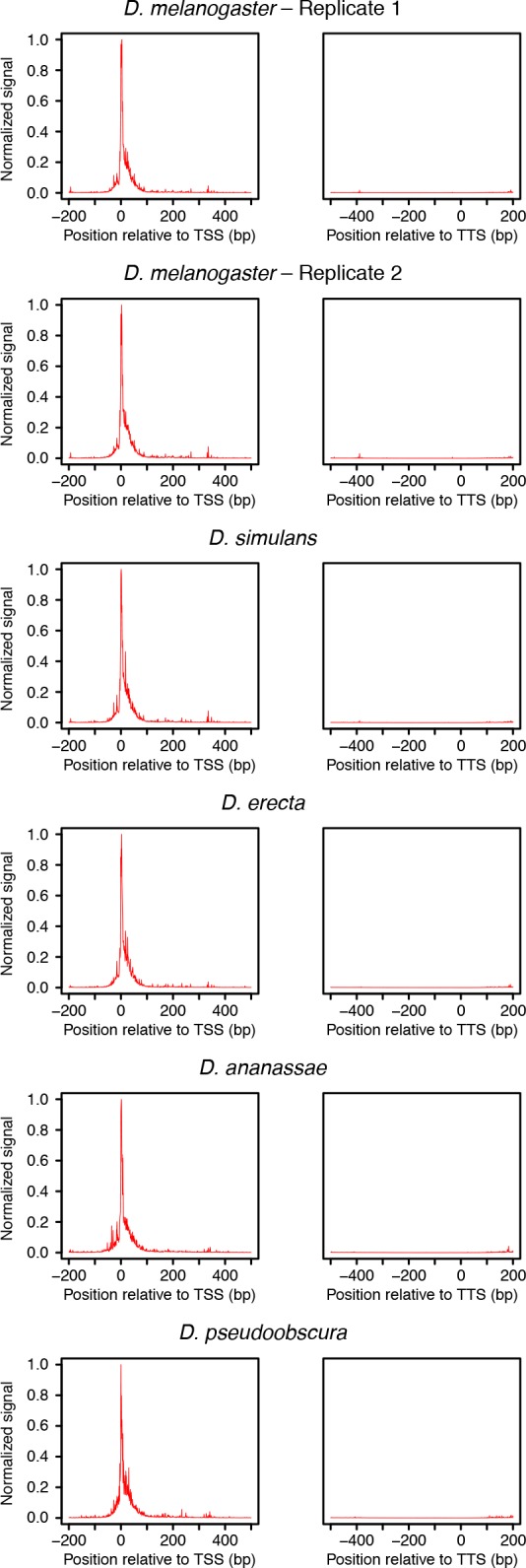
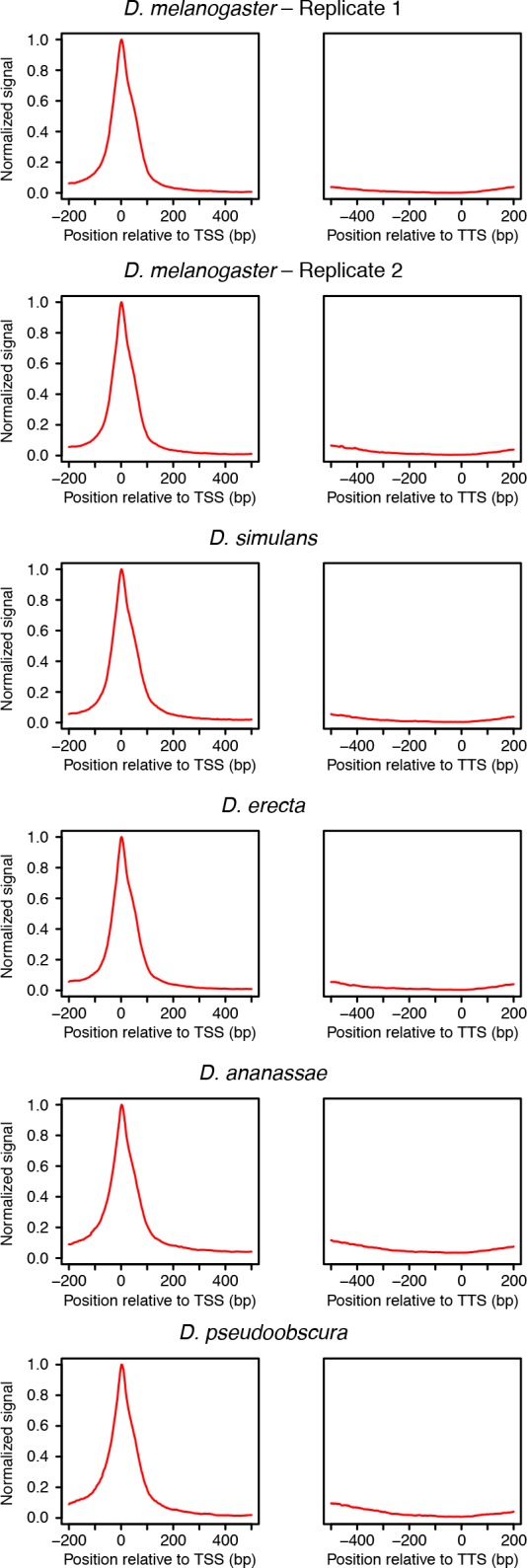
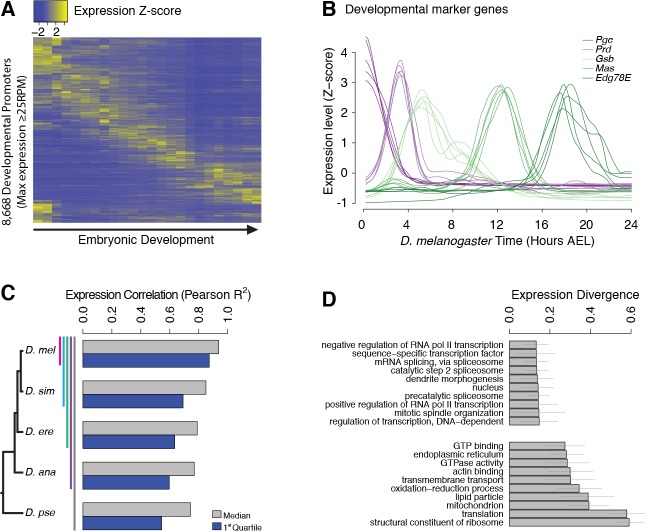
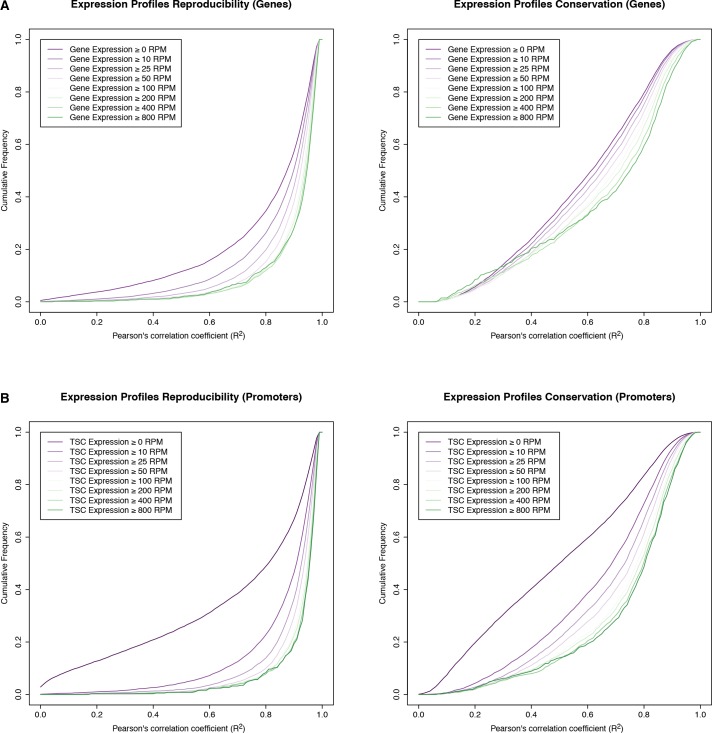
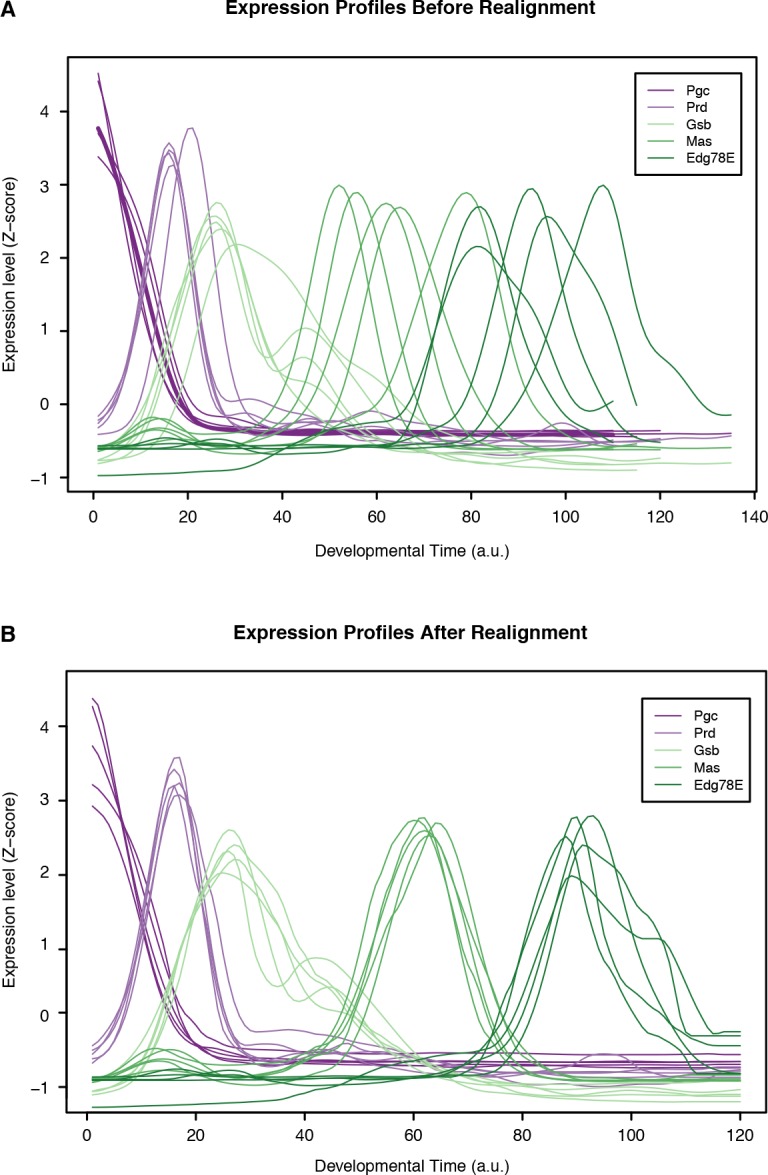
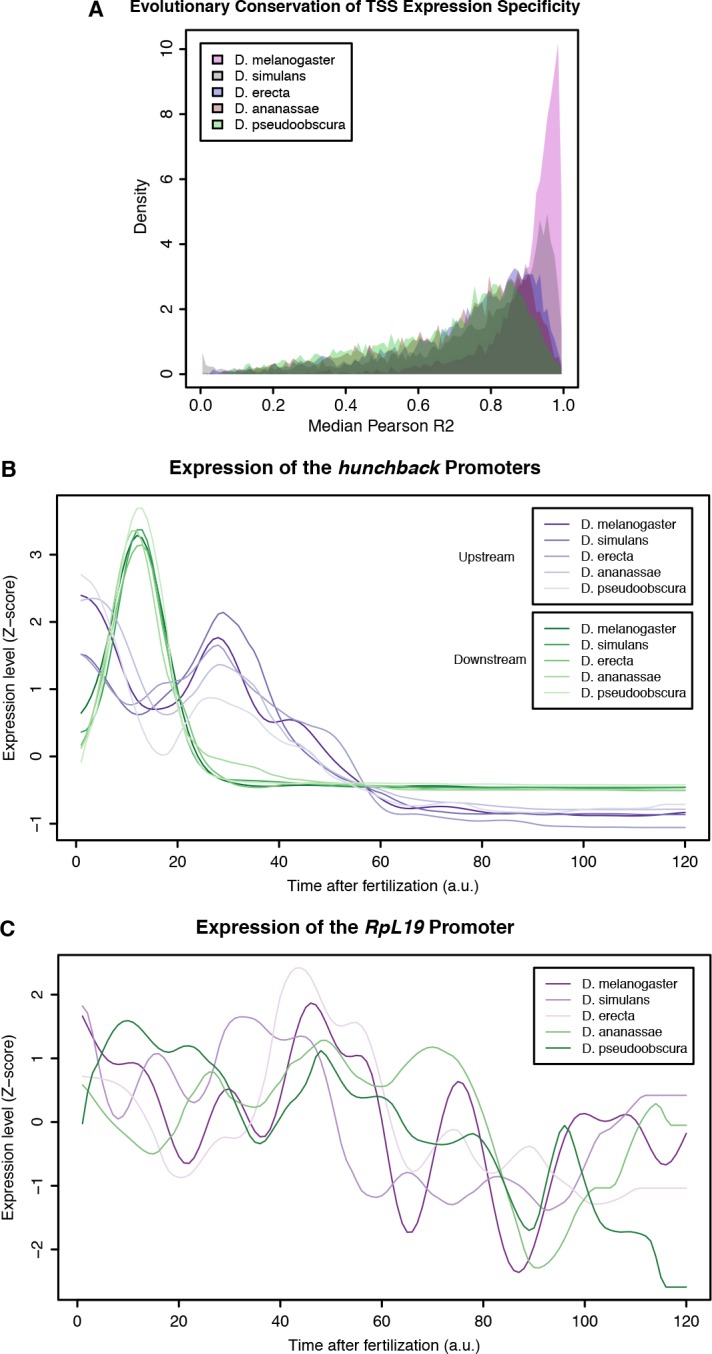
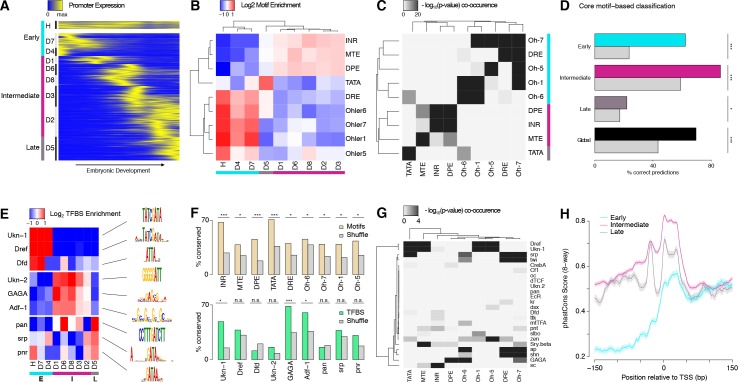
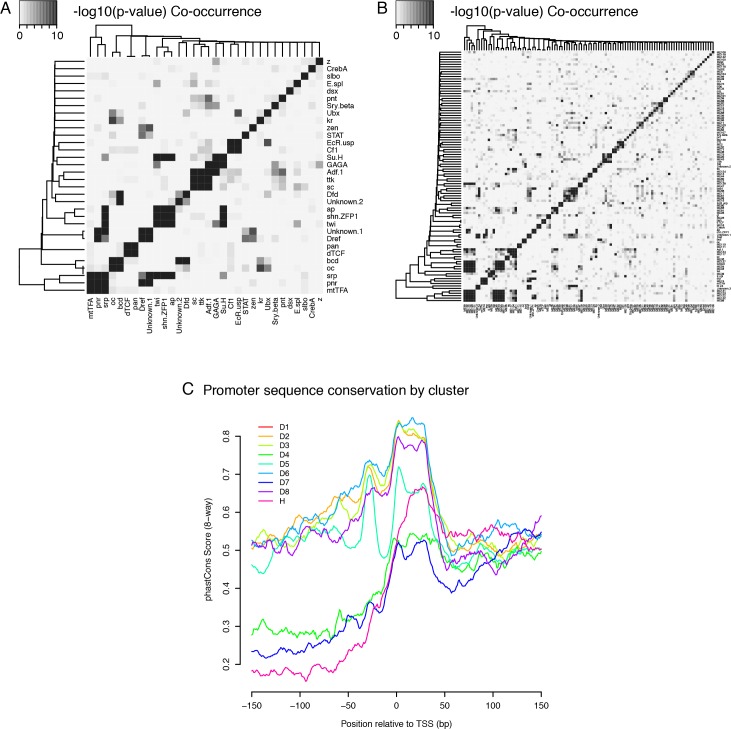
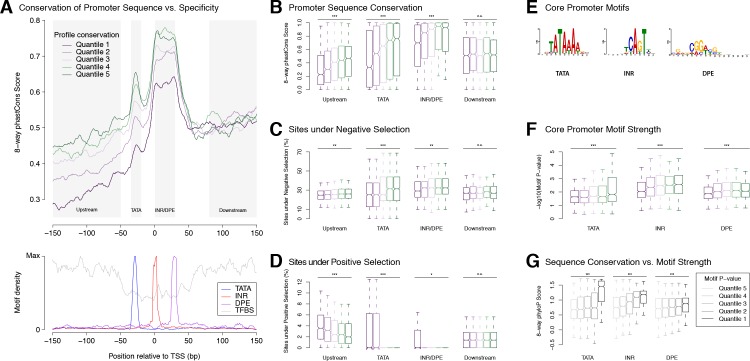
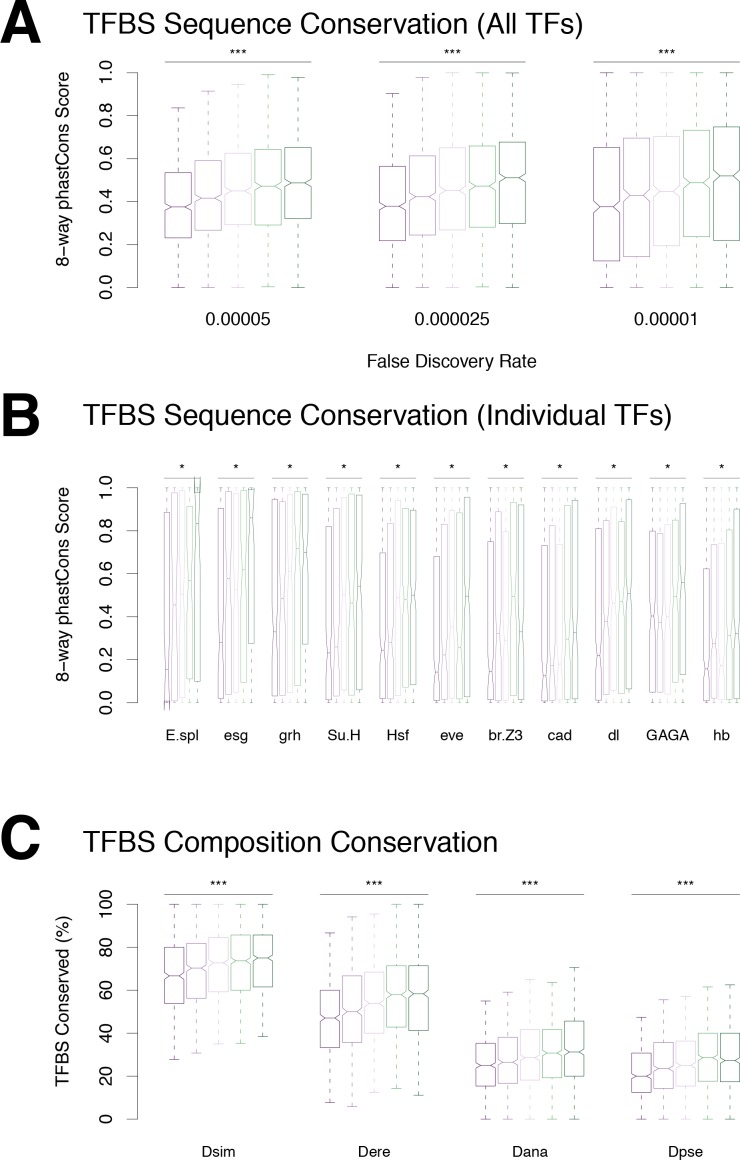
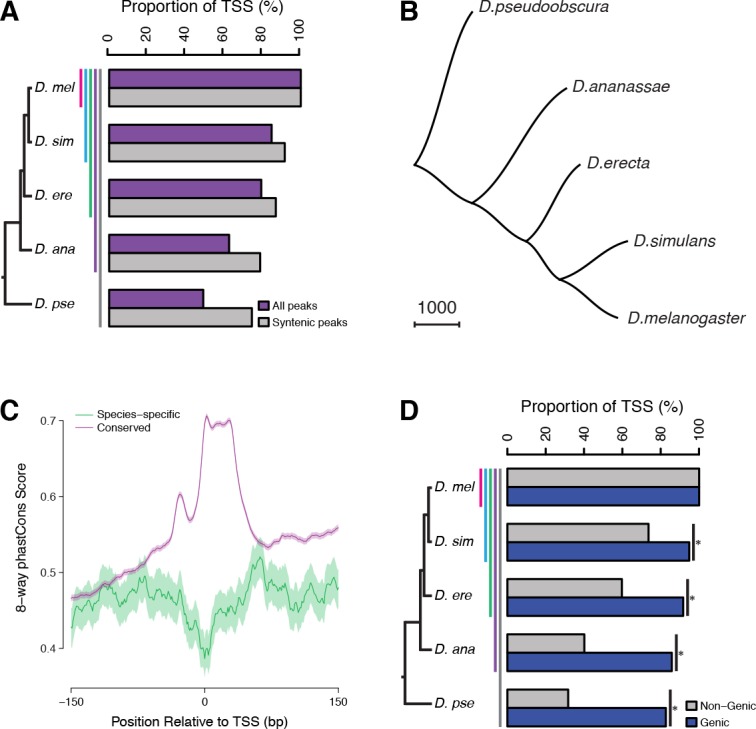
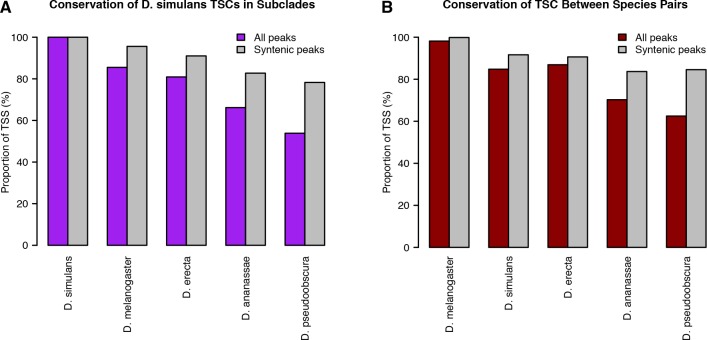
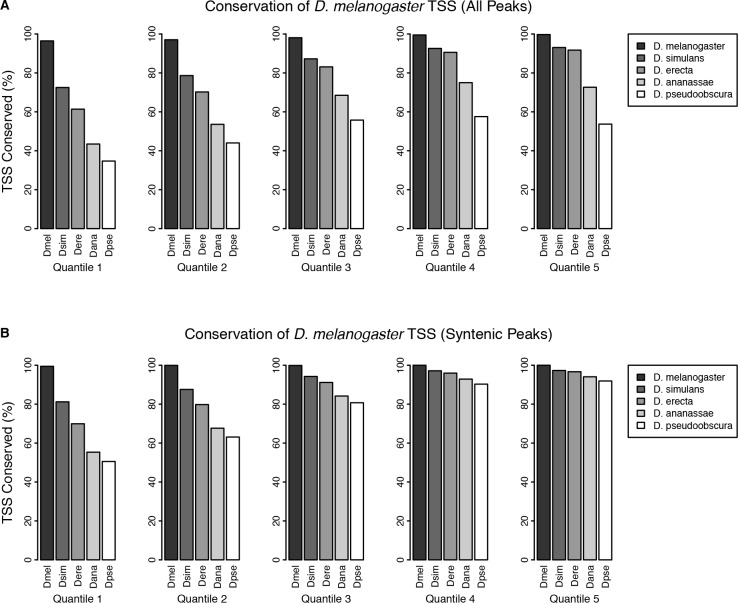
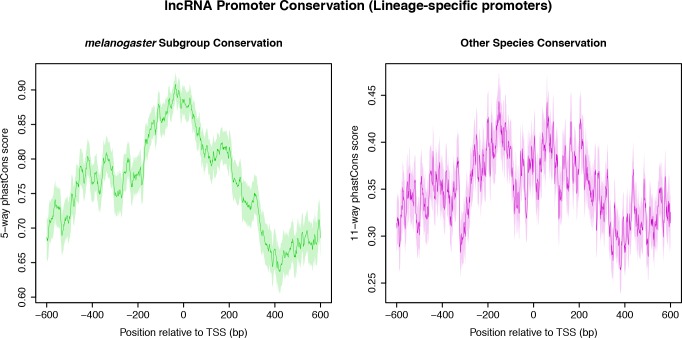
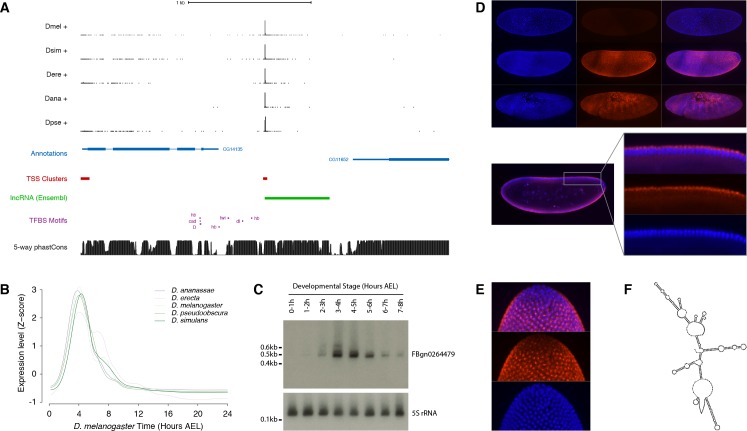
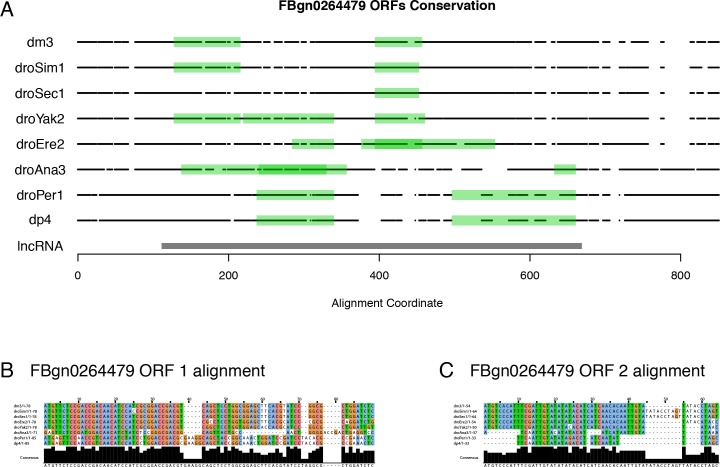
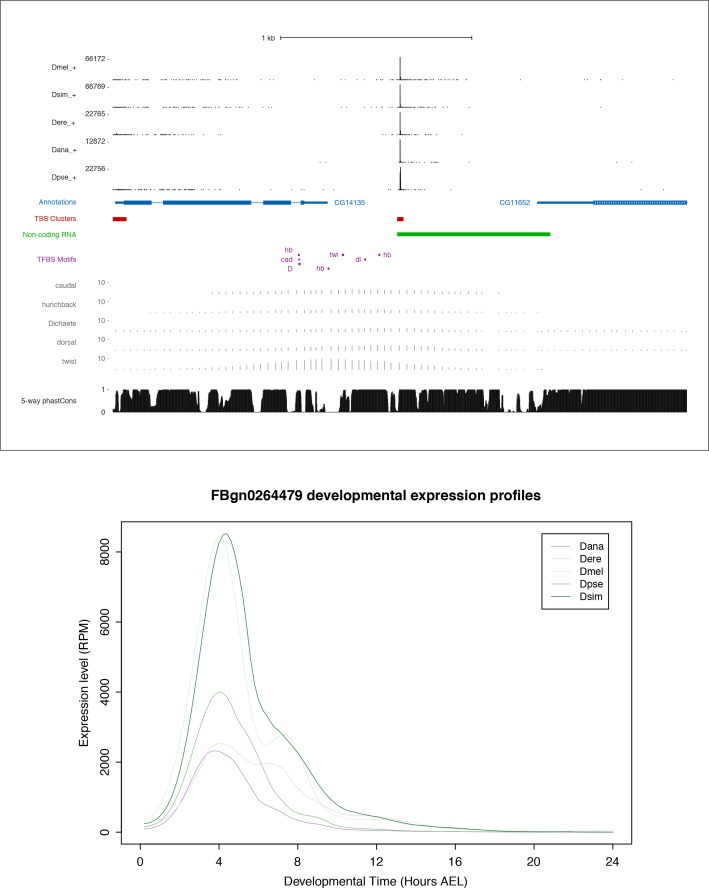
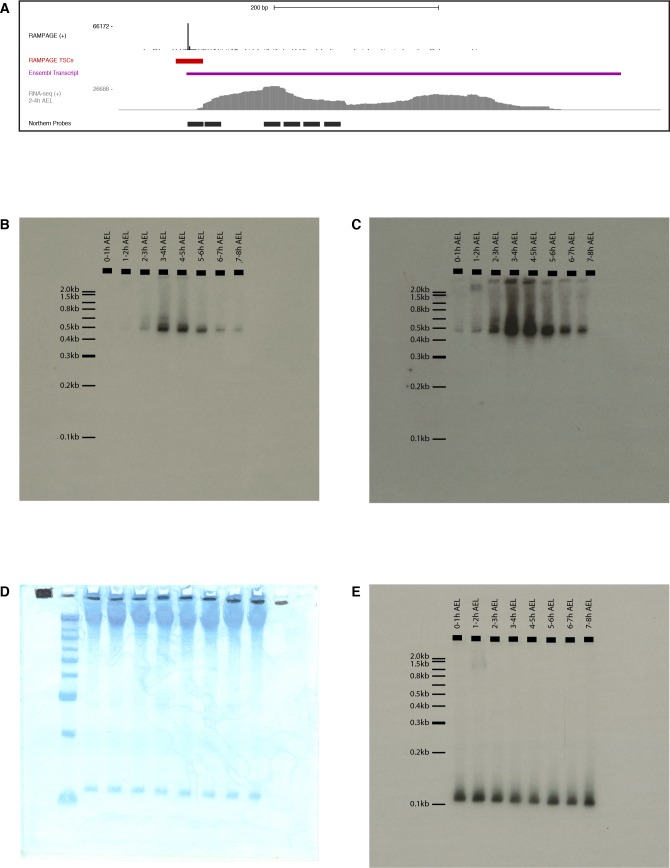
Multicellular development is driven by regulatory programs that orchestrate the transcription of protein-coding and noncoding genes. To decipher this genomic regulatory code, and to investigate the developmental relevance of noncoding transcription, we compared genome-wide promoter activity throughout embryogenesis in 5 species. Core promoters, generally not thought to play a significant regulatory role, in fact impart restrictions on the developmental timing of gene expression on a global scale. We propose a hierarchical regulatory model in which core promoters define broad windows of opportunity for expression, by defining a range of transcription factors from which they can receive regulatory inputs. This two-tiered mechanism globally orchestrates developmental gene expression, including extremely widespread noncoding transcription. The sequence and expression specificity of noncoding RNA promoters are evolutionarily conserved, implying biological relevance. Overall, this work introduces a hierarchical model for developmental gene regulation, and reveals a major role for noncoding transcription in animal development.
多细胞发育是由调节程序驱动的,这些程序协调蛋白质编码和非编码基因的转录。为了解读这个基因组调控密码,并研究非编码转录的发育相关性,我们在 5 个物种的胚胎发生过程中比较了全基因组启动子活性。核心启动子通常被认为没有重要的调控作用,但实际上在全局范围内对基因表达的发育时间施加了限制。我们提出了一个层次化的调控模型,其中核心启动子通过定义一个转录因子范围来定义表达的机会窗口,从而可以接收调节输入。这种两层机制在全球范围内协调发育基因表达,包括极其广泛的非编码转录。非编码 RNA 启动子的序列和表达特异性在进化上是保守的,暗示了其具有生物学相关性。总的来说,这项工作介绍了一种发育基因调控的层次模型,并揭示了非编码转录在动物发育中的主要作用。