Masschelein Joleen, Clauwers Charlien, Awodi Ufedo R, Stalmans Karen, Vermaelen Wesley, Lescrinier Eveline, Aertsen Abram, Michiels Chris, Challis Gregory L, Lavigne Rob
Laboratory of Gene Technology , KU Leuven , Kasteelpark Arenberg 21-box 2462 , B-3001 Heverlee , Belgium . Email:
Laboratory of Food Microbiology , KU Leuven , Kasteelpark Arenberg 22 , B-3001 Heverlee , Belgium.
Chem Sci. 2015 Feb 1;6(2):923-929. doi: 10.1039/c4sc01927j. Epub 2014 Oct 15.
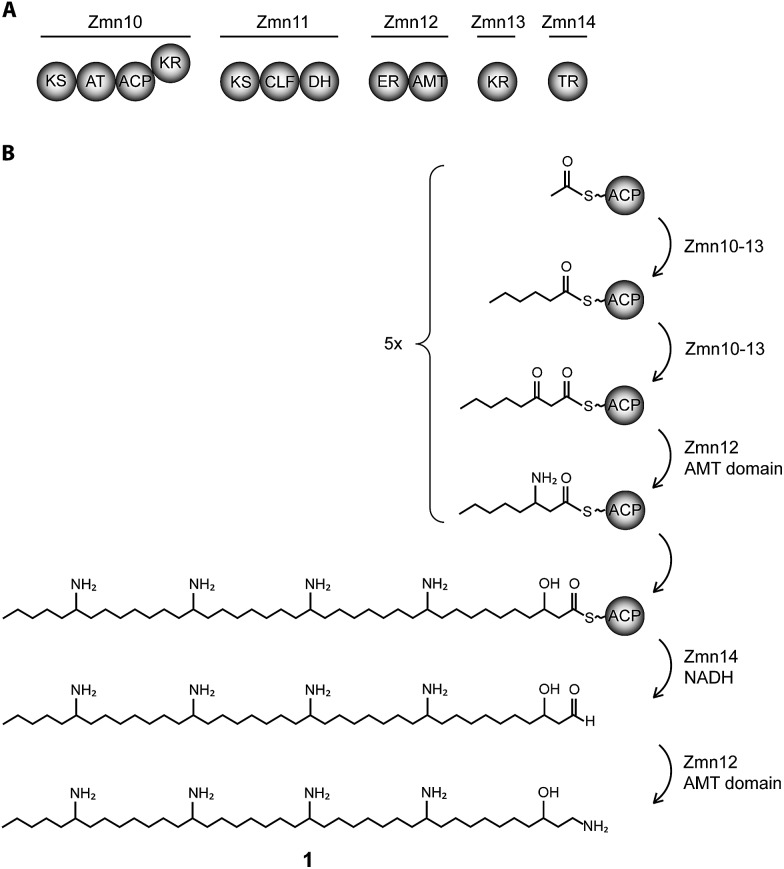
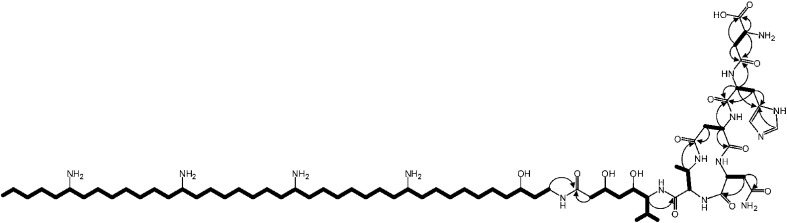
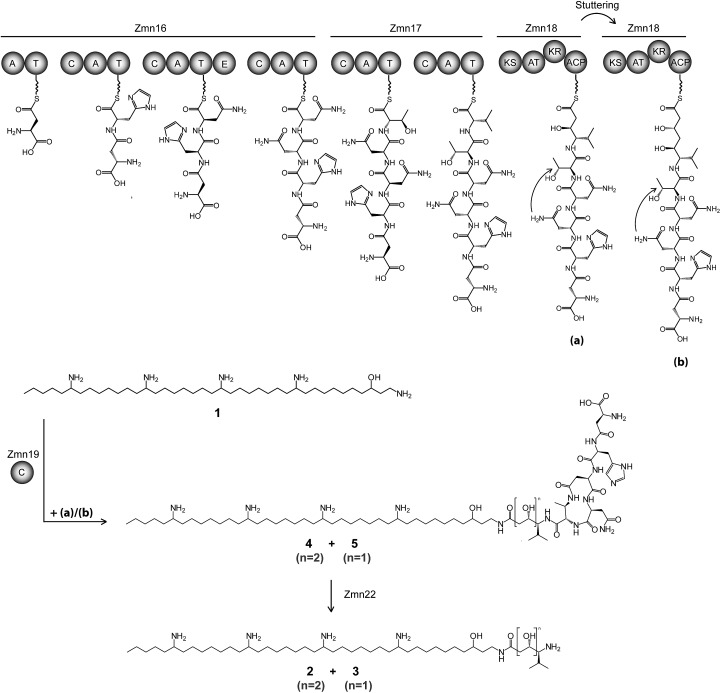
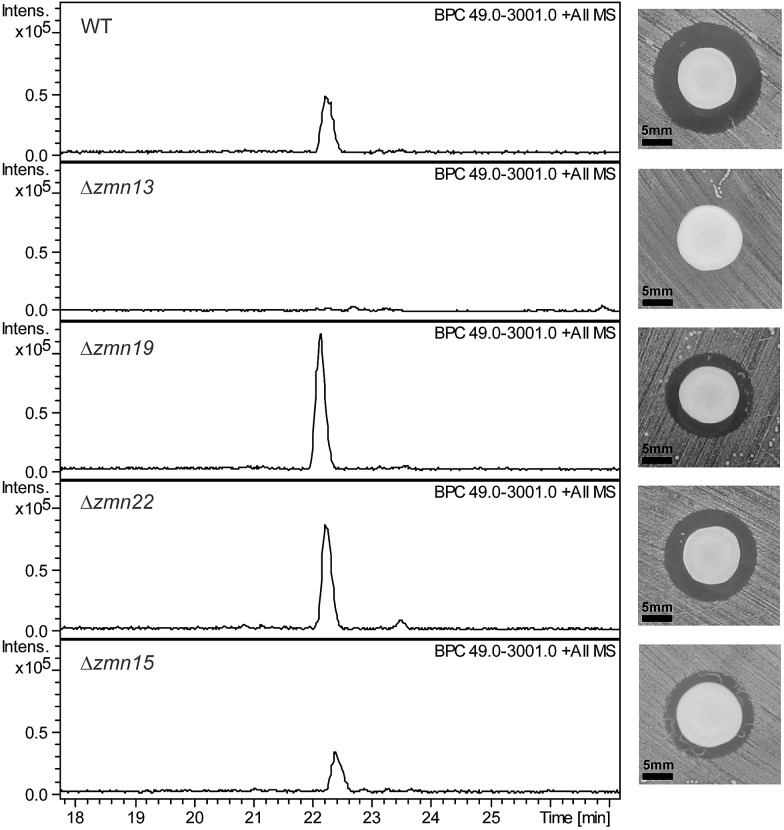
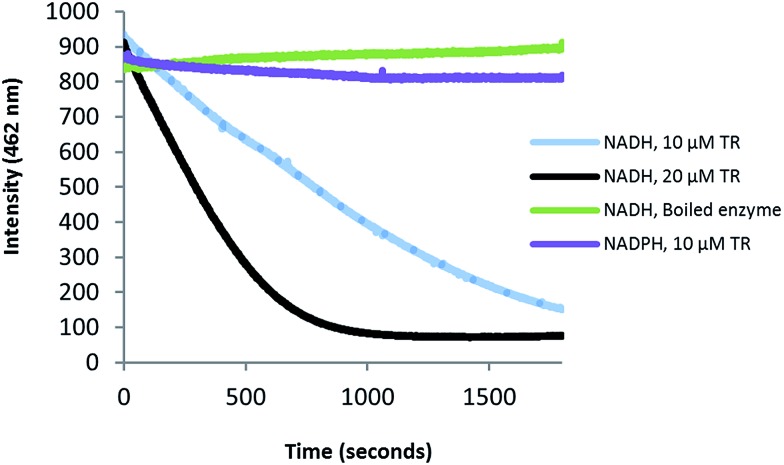
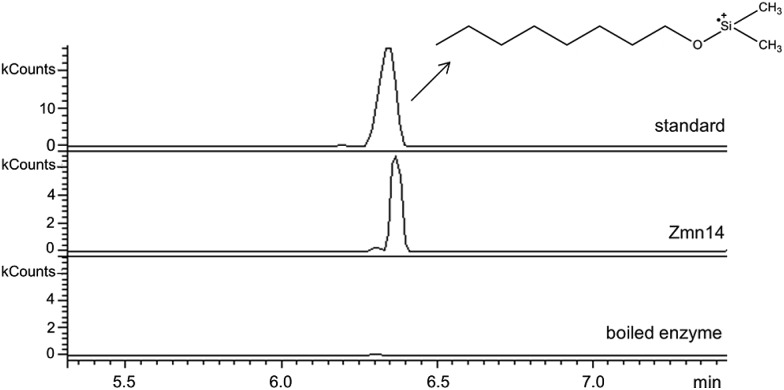
The zeamines are a unique group of antibiotics produced by RVH1 that contain variable hybrid peptide-polyketide moieties connected to a common pentaamino-hydroxyalkyl chain. They exhibit potent activity against a broad spectrum of Gram-positive and Gram-negative bacteria. Here we report a combination of targeted gene deletions, high resolution LC-MS(/MS) analyses, biochemical assays and feeding studies that define the functions of several key zeamine biosynthetic enzymes. The pentaamino-hydroxyalkyl chain is assembled by an iterative multienzyme complex (Zmn10-13) that bears a close resemblance to polyunsaturated fatty acid synthases. Zmn14 was shown to function as an NADH-dependent thioester reductase and is proposed to release a tetraamino-hydroxyalkyl thioester from the acyl carrier protein domain of Zmn10 as an aldehyde. Despite the intrinsic ability of Zmn14 to catalyze further reduction of aldehydes to alcohols, the initially-formed aldehyde intermediate is proposed to undergo preferential transamination to produce zeamine II. In a parallel pathway, hexapeptide-monoketide and hexapeptide-diketide thioesters are generated by a hybrid nonribosomal peptide synthetase-polyketide synthase multienzyme complex (Zmn16-18) and subsequently conjugated to zeamine II by a stand-alone condensing enzyme (Zmn19). Structures for the resulting prezeamines were elucidated using a combination of high resolution LC-MS/MS and 1- and 2-D NMR spectroscopic analyses. The prezeamines are hypothesized to be precursors of the previously-identified zeamines, which are generated by the action of Zmn22, an acylpeptide hydrolase that specifically cleaves the N-terminal pentapeptide of the prezeamines in a post-assembly processing step. Thus, the zeamine antibiotics are assembled by a unique combination of nonribosomal peptide synthetase, type I modular polyketide synthase and polyunsaturated fatty acid synthase-like biosynthetic machinery.
玉米赤霉烯胺是由RVH1产生的一类独特抗生素,其包含与一条共同的五氨基 - 羟烷基链相连的可变杂合肽 - 聚酮部分。它们对多种革兰氏阳性菌和革兰氏阴性菌具有强效活性。在此,我们报告了一系列靶向基因缺失、高分辨率液相色谱 - 质谱联用(/串联质谱)分析、生化测定和饲喂研究,这些研究确定了几种关键玉米赤霉烯胺生物合成酶的功能。五氨基 - 羟烷基链由一个与多不饱和脂肪酸合酶极为相似的迭代多酶复合物(Zmn10 - 13)组装而成。Zmn14被证明作为一种依赖NADH的硫酯还原酶发挥作用,并被认为从Zmn10的酰基载体蛋白结构域释放出一种四氨基 - 羟烷基硫酯作为醛。尽管Zmn14具有将醛进一步还原为醇的内在能力,但最初形成的醛中间体被认为优先进行转氨作用以产生玉米赤霉烯胺II。在一条平行途径中,六肽 - 单酮和六肽 - 二酮硫酯由一个杂合的非核糖体肽合成酶 - 聚酮合成酶多酶复合物(Zmn16 - 18)产生,随后通过一种独立的缩合酶(Zmn19)与玉米赤霉烯胺II共轭。使用高分辨率液相色谱 - 串联质谱以及一维和二维核磁共振光谱分析相结合的方法阐明了所得前玉米赤霉烯胺的结构。前玉米赤霉烯胺被假定为先前鉴定的玉米赤霉烯胺的前体,后者是由Zmn22(一种酰基肽水解酶)的作用产生的,该酶在组装后加工步骤中特异性切割前玉米赤霉烯胺的N端五肽。因此,玉米赤霉烯胺类抗生素是由非核糖体肽合成酶、I型模块化聚酮合成酶和多不饱和脂肪酸合酶样生物合成机制的独特组合组装而成。