Systems Biology Program, Centre for Genomic Regulation, The Barcelona Institute for Science and Technology, Barcelona, Spain.
Universitat Pompeu Fabra, Barcelona, Spain.
Elife. 2018 Apr 11;7:e32472. doi: 10.7554/eLife.32472.
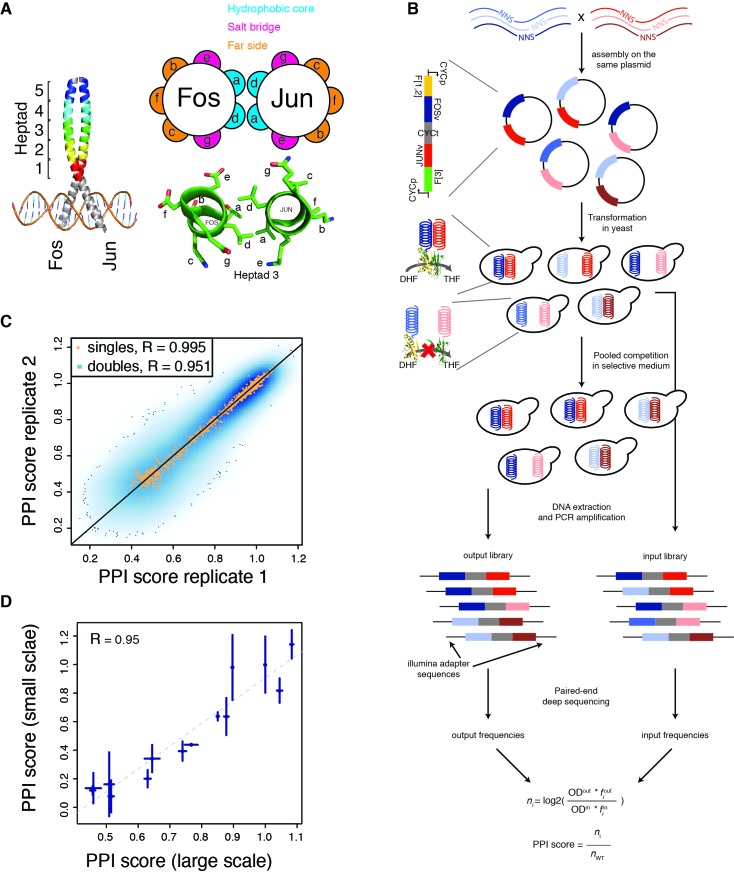
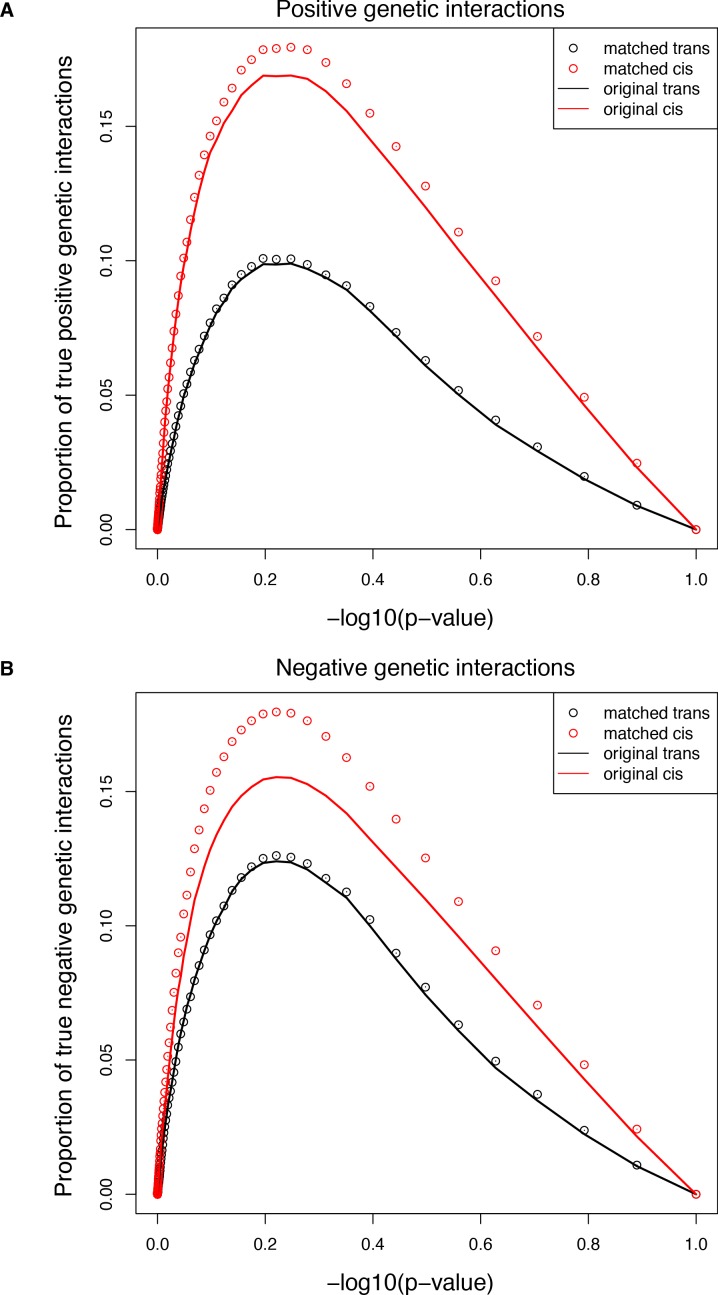
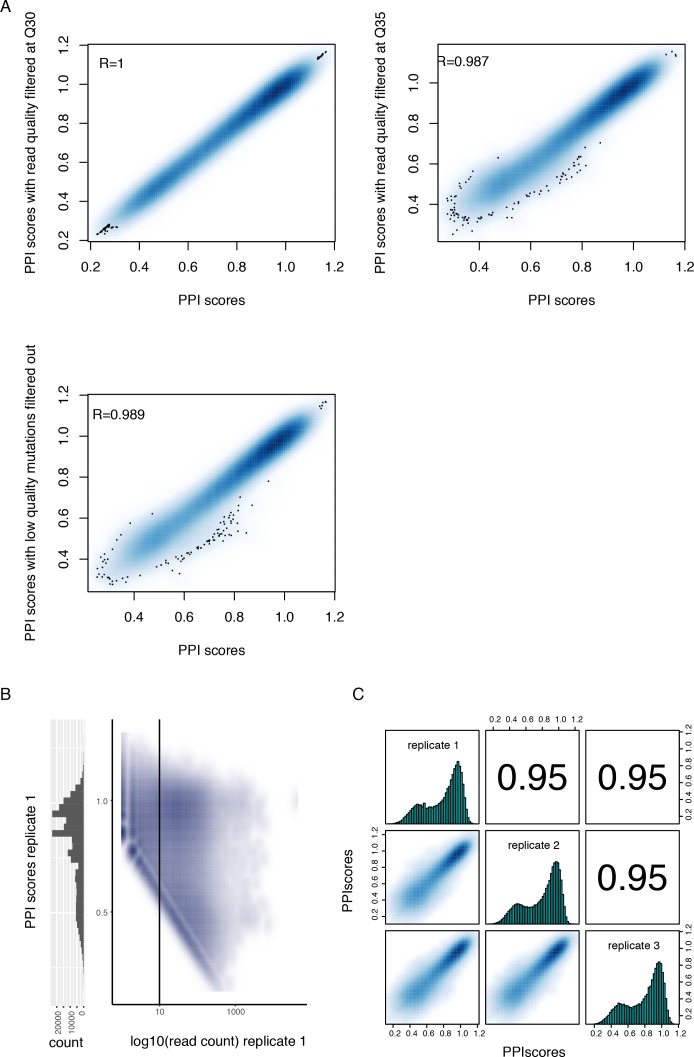
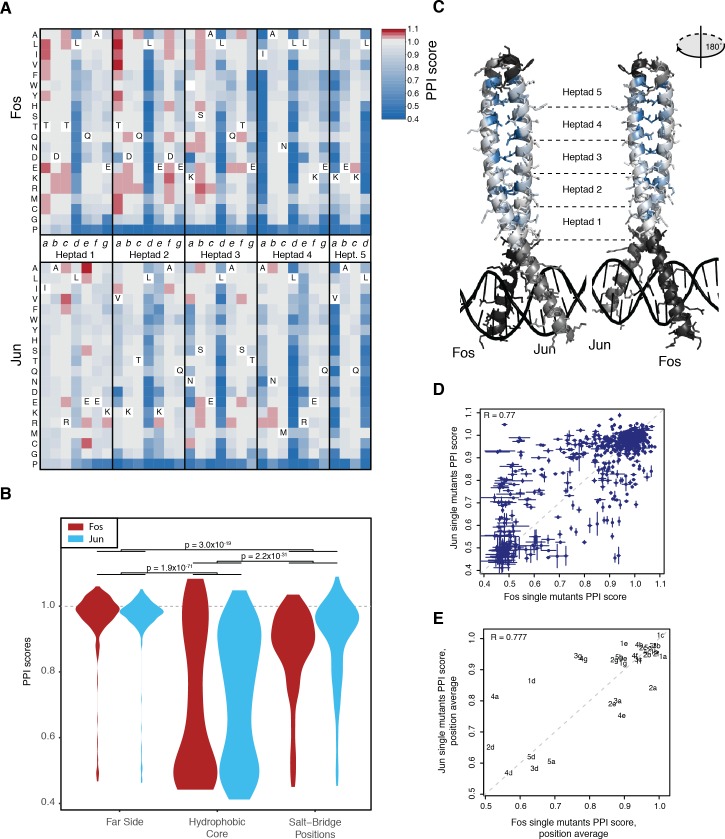
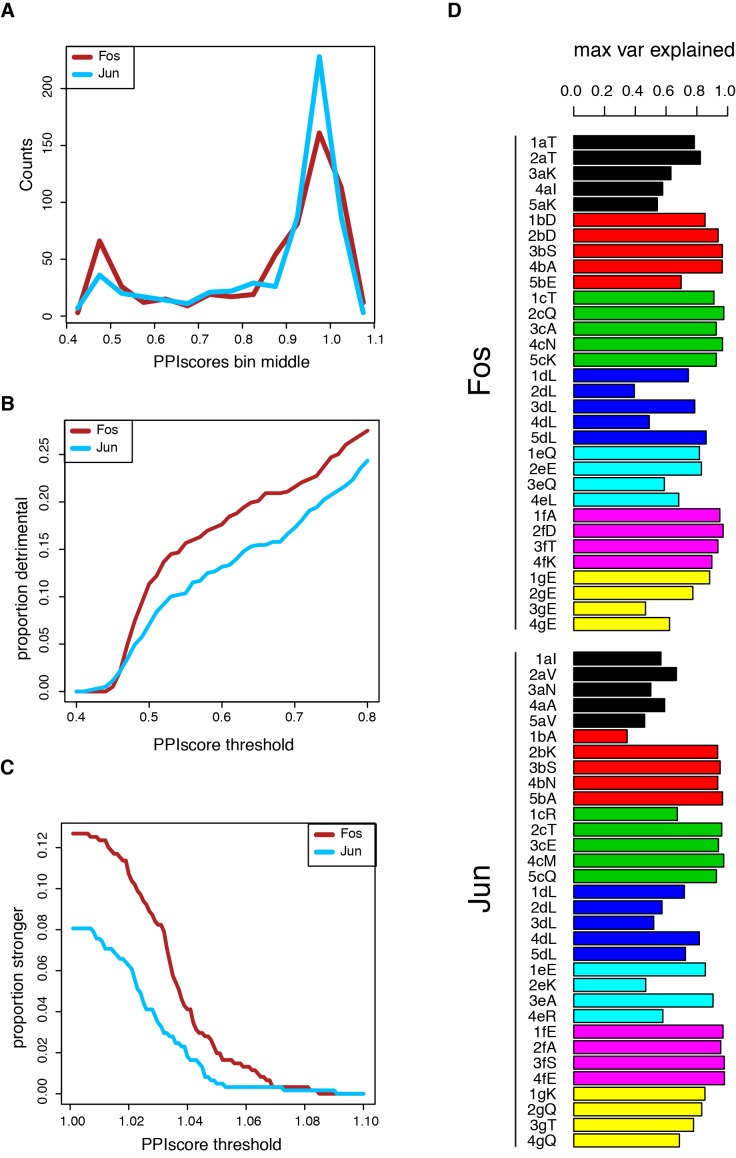
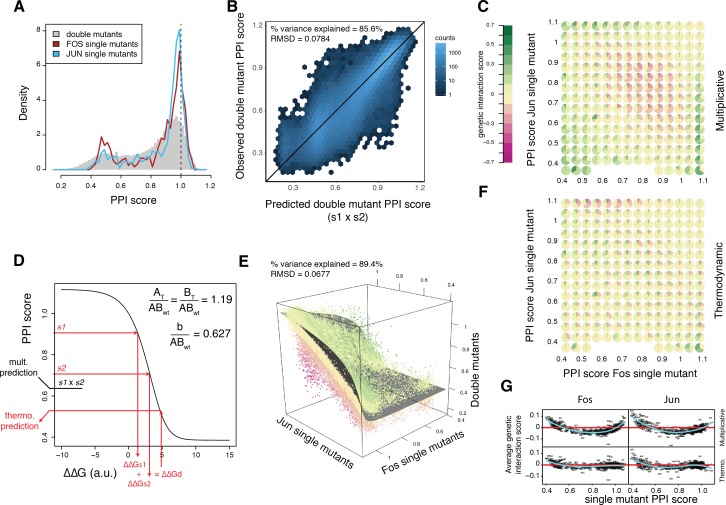
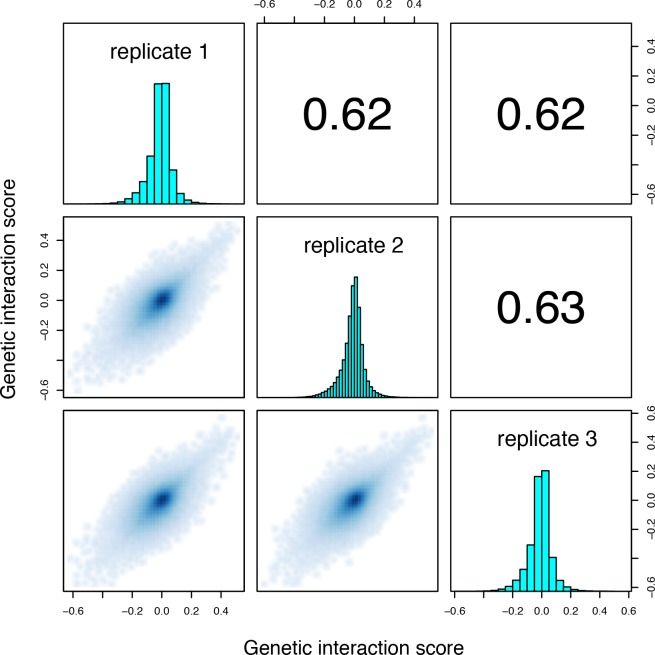
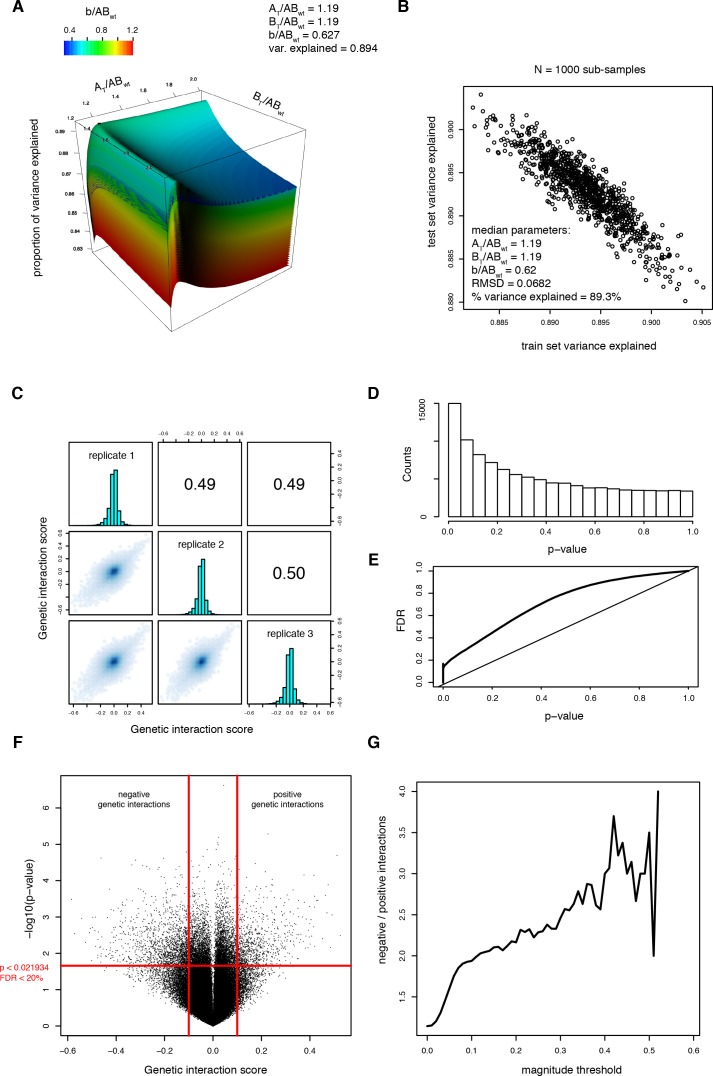
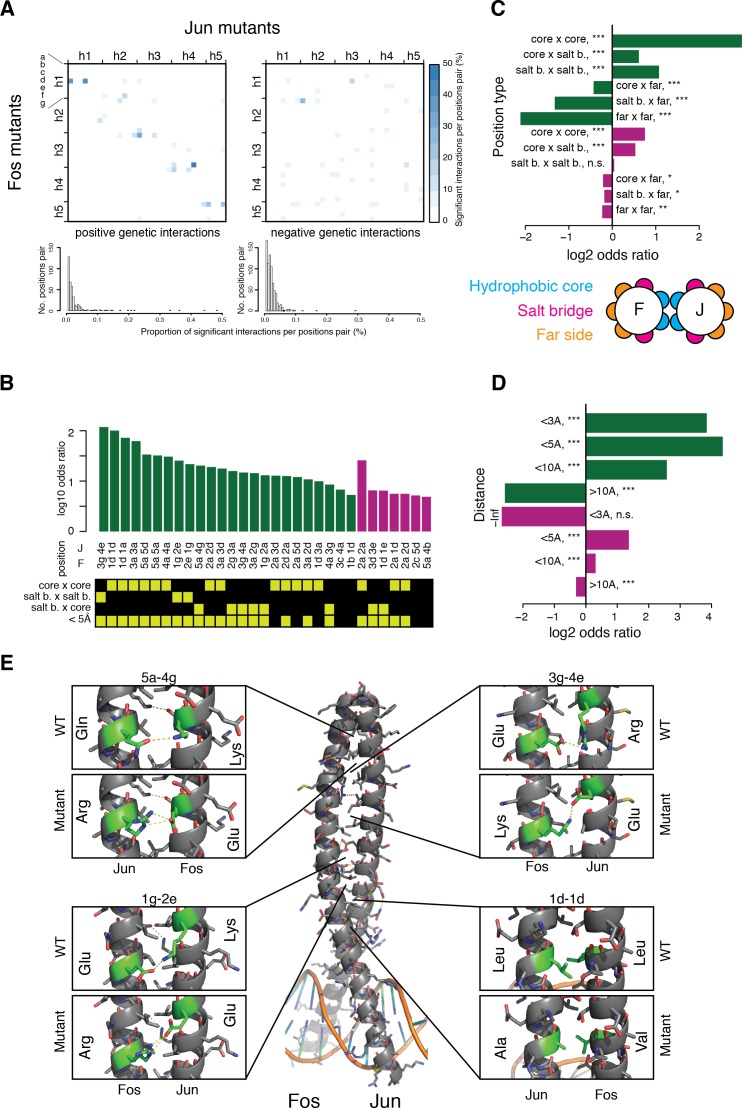
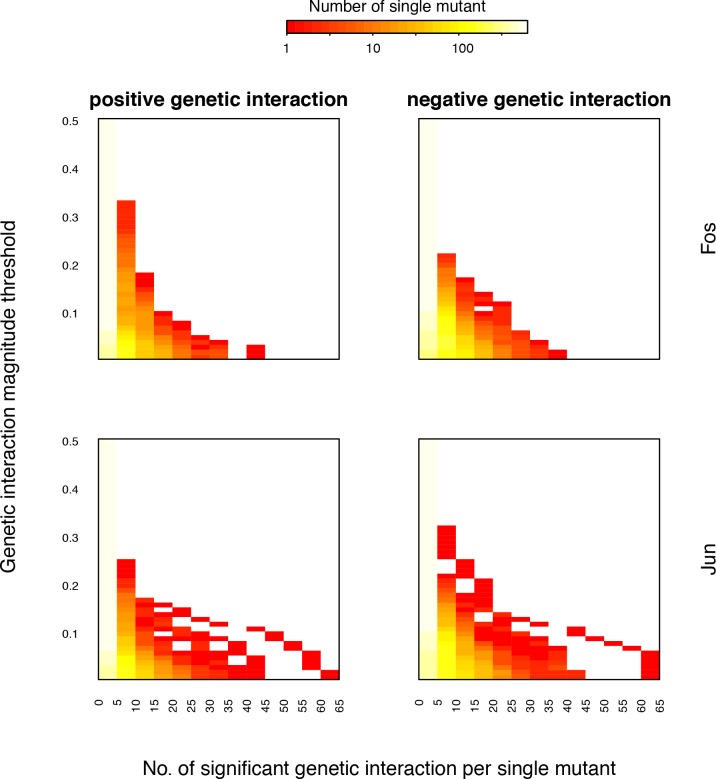
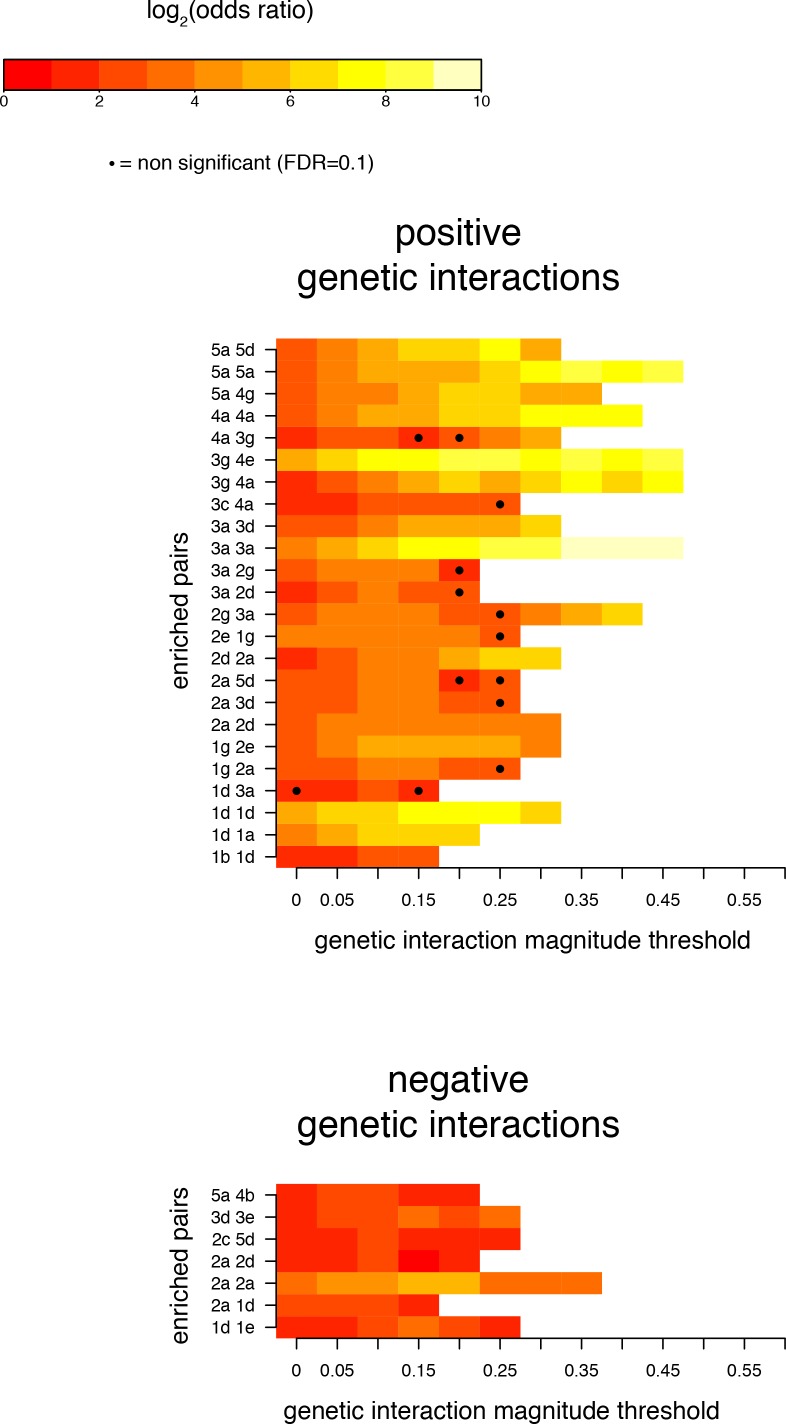
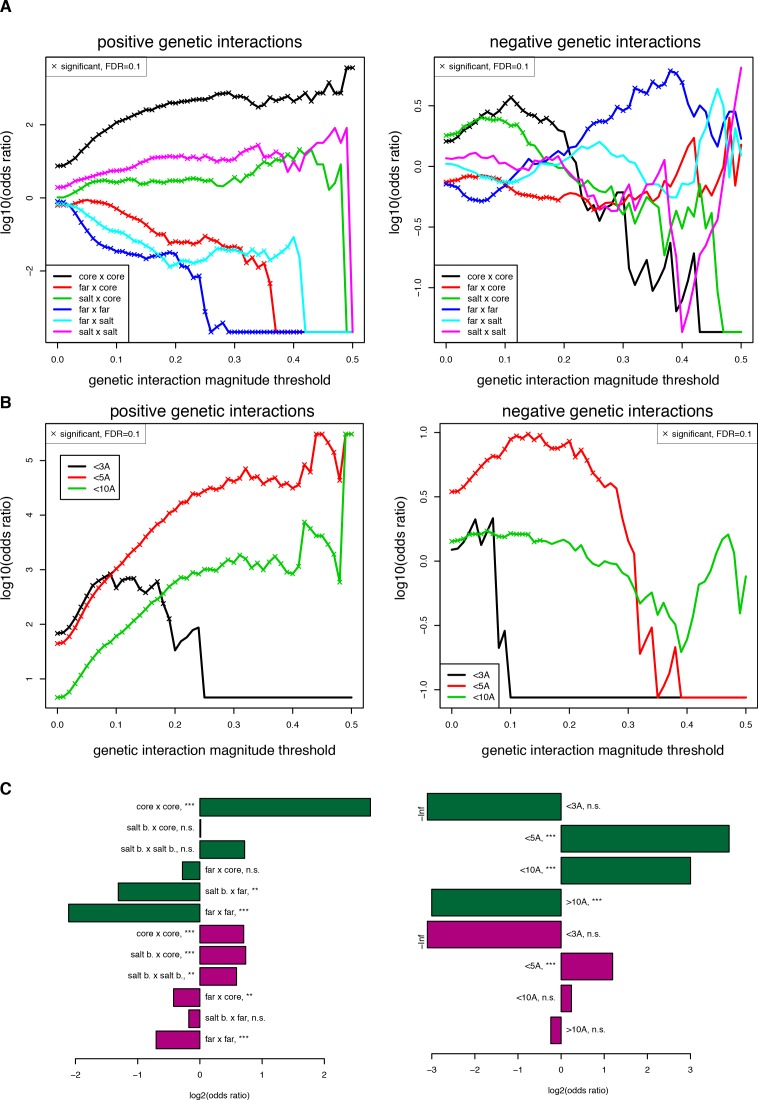
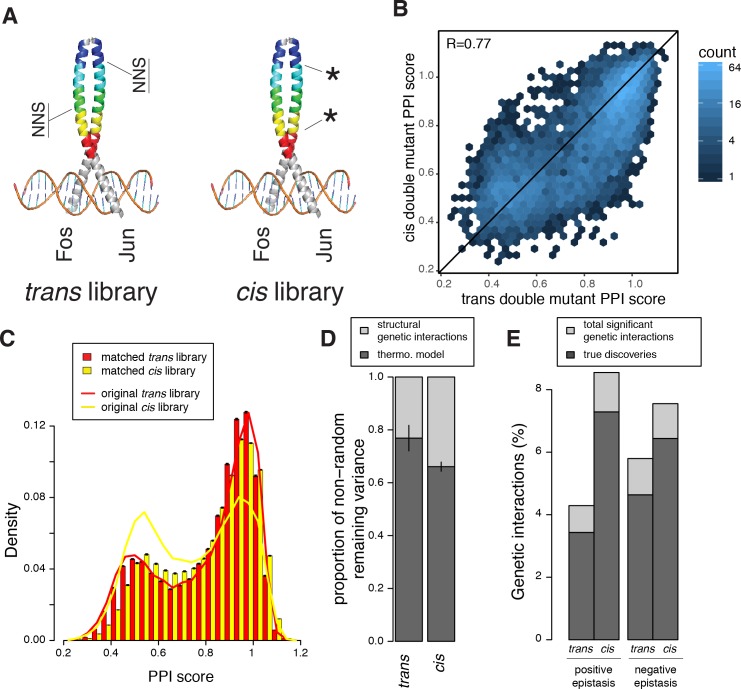
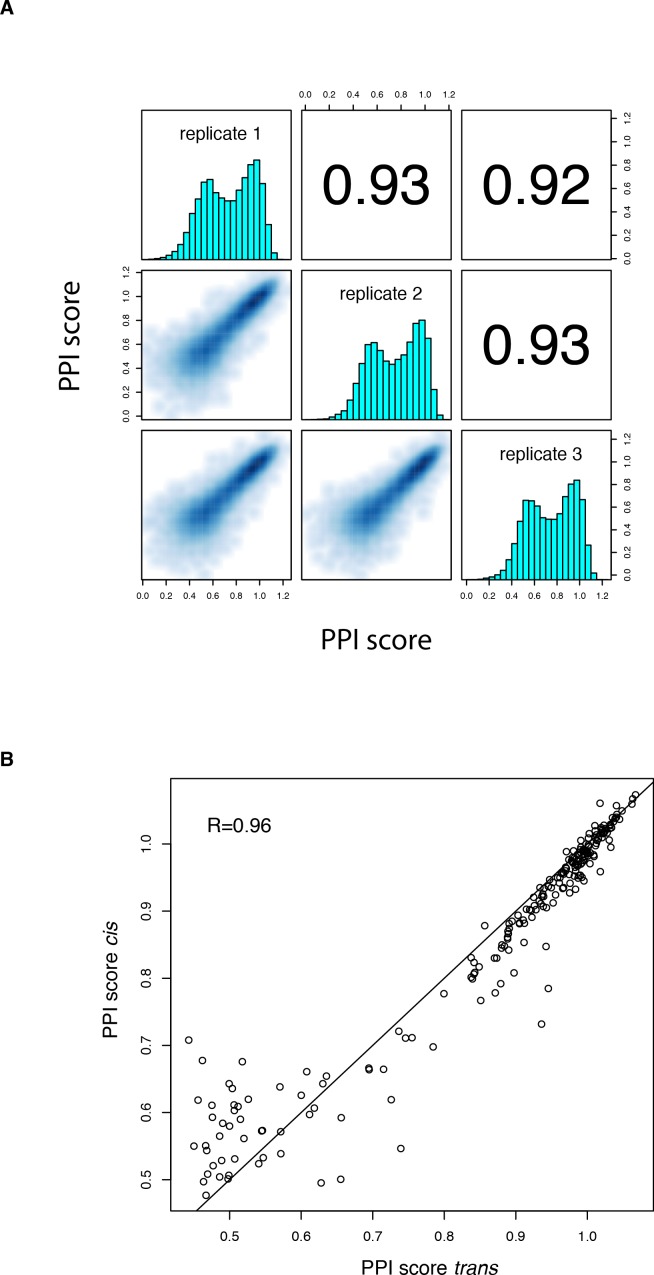
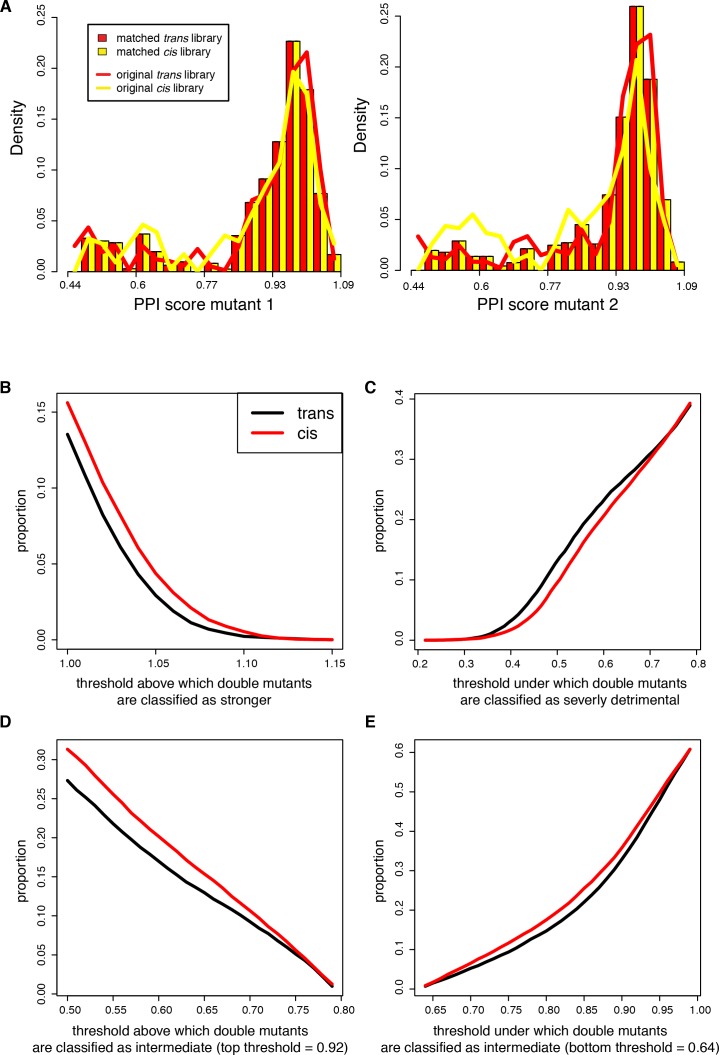
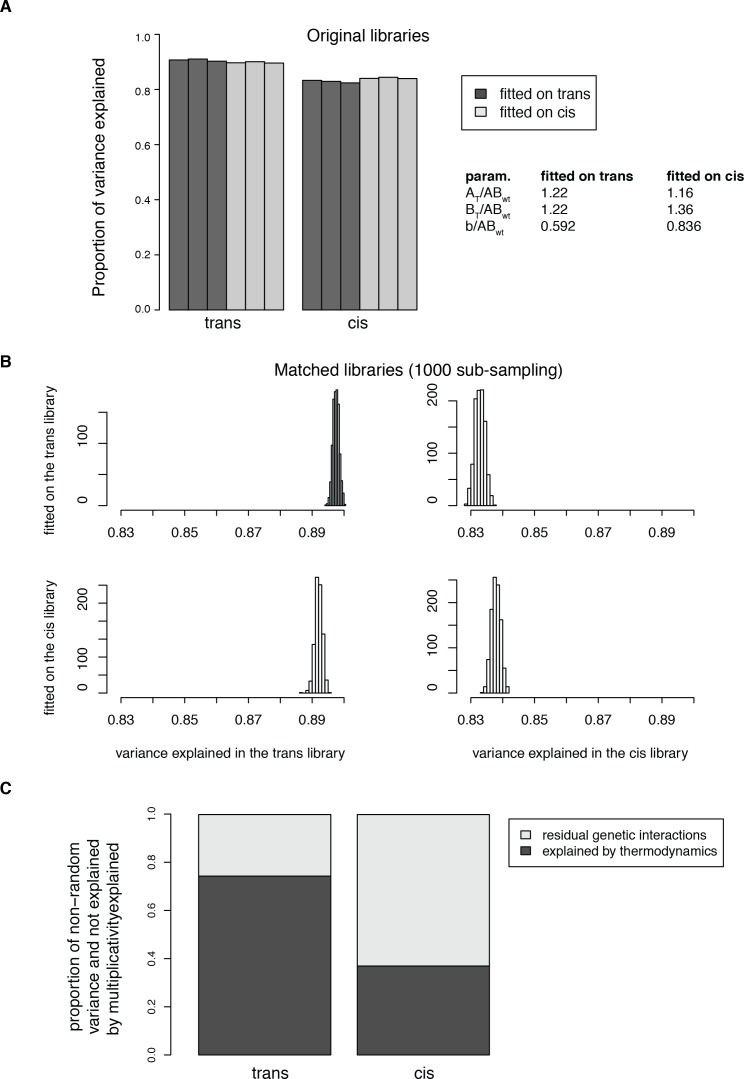
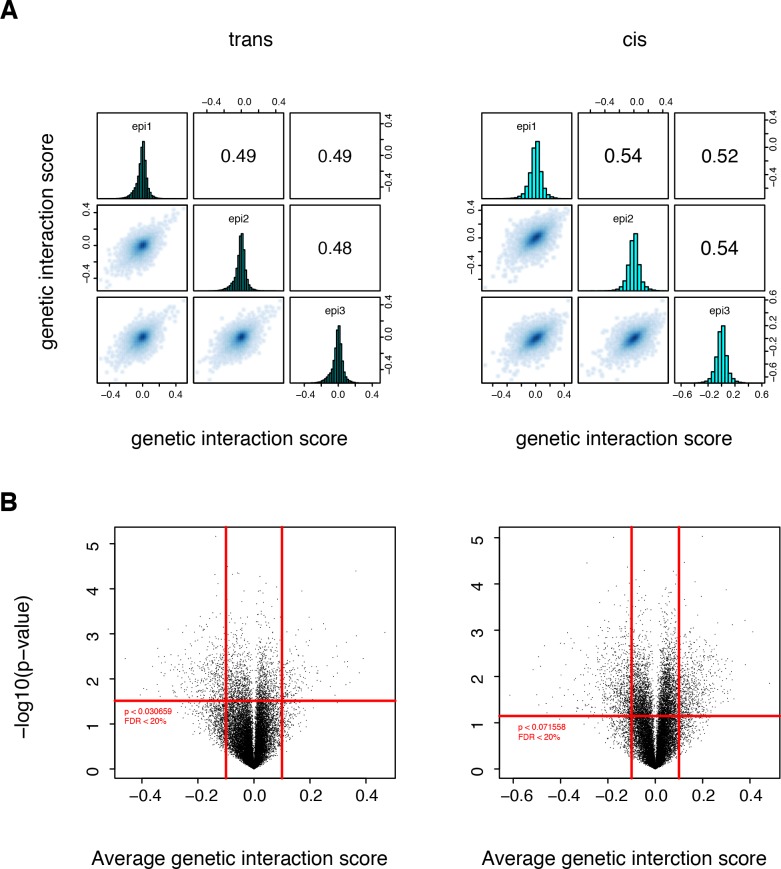
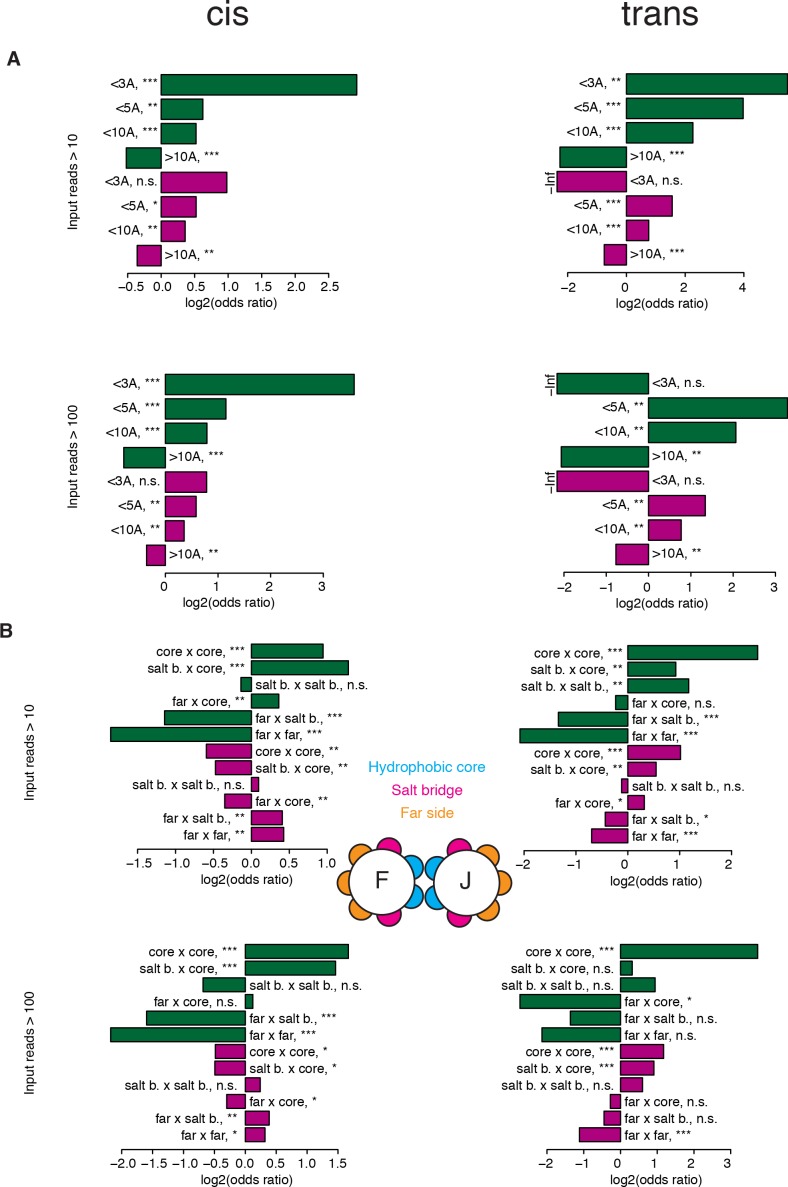
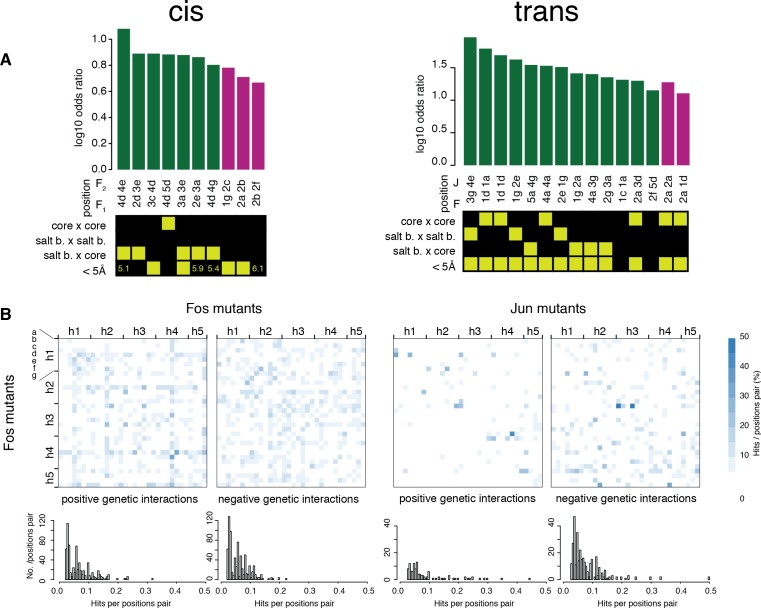
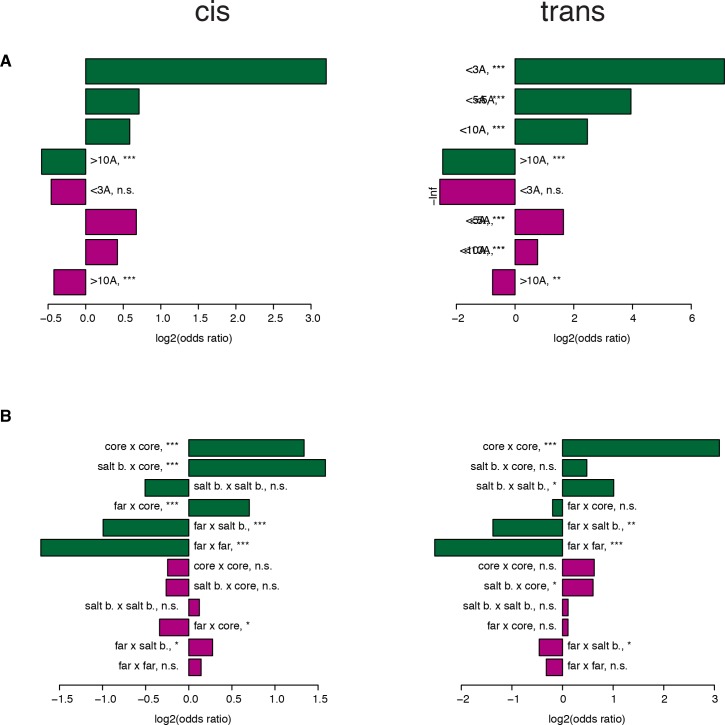
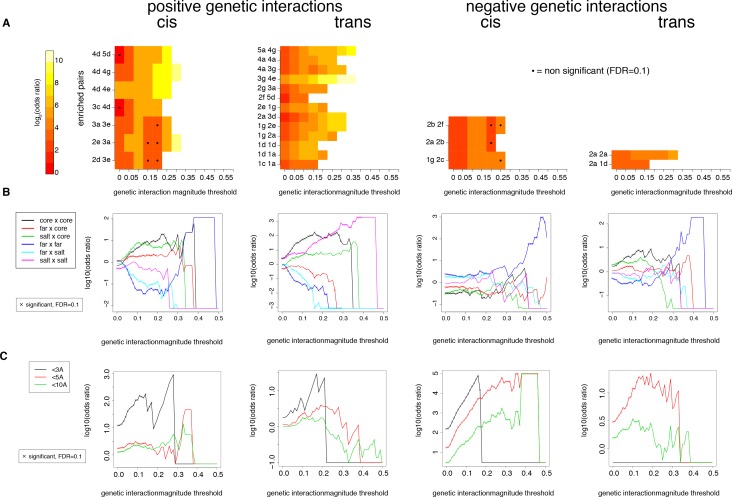
A key question in human genetics and evolutionary biology is how mutations in different genes combine to alter phenotypes. Efforts to systematically map genetic interactions have mostly made use of gene deletions. However, most genetic variation consists of point mutations of diverse and difficult to predict effects. Here, by developing a new sequencing-based protein interaction assay - - we quantified the effects of >120,000 pairs of point mutations on the formation of the AP-1 transcription factor complex between the products of the FOS and JUN proto-oncogenes. Genetic interactions are abundant both in (within one protein) and (between the two molecules) and consist of two classes - interactions driven by thermodynamics that can be predicted using a three-parameter global model, and structural interactions between proximally located residues. These results reveal how physical interactions generate quantitatively predictable genetic interactions.
在人类遗传学和进化生物学中,一个关键问题是不同基因中的突变如何组合改变表型。系统绘制遗传相互作用的努力主要利用了基因缺失。然而,大多数遗传变异由点突变组成,其影响多样且难以预测。在这里,通过开发一种新的基于测序的蛋白质相互作用测定法 - ,我们定量测定了超过 120,000 对点突变对 FOS 和 JUN 原癌基因产物之间形成 AP-1 转录因子复合物的影响。遗传相互作用在 (一个蛋白质内)和 (两个分子之间)都很丰富,由两类组成 - 由热力学驱动的相互作用,可以使用三参数全局模型进行预测,以及近距离残基之间的结构相互作用。这些结果揭示了物理相互作用如何产生可定量预测的遗传相互作用。