Telomere Biology Section, Laboratory of Biochemistry and Molecular Biology, National Cancer Institute, National Institutes of Health, Bethesda, Maryland 20892, USA.
Telomere Biology Laboratory, Cancer Research UK, London Research Institute, London WC2A 3LY, United Kingdom.
Genes Dev. 2018 Apr 1;32(7-8):537-554. doi: 10.1101/gad.311712.118. Epub 2018 Apr 13.
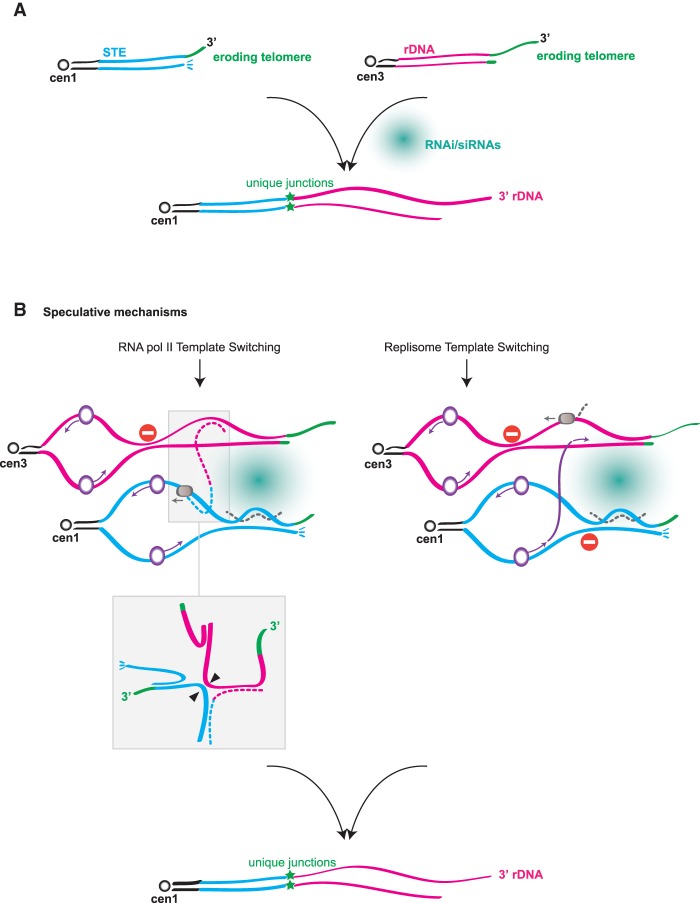
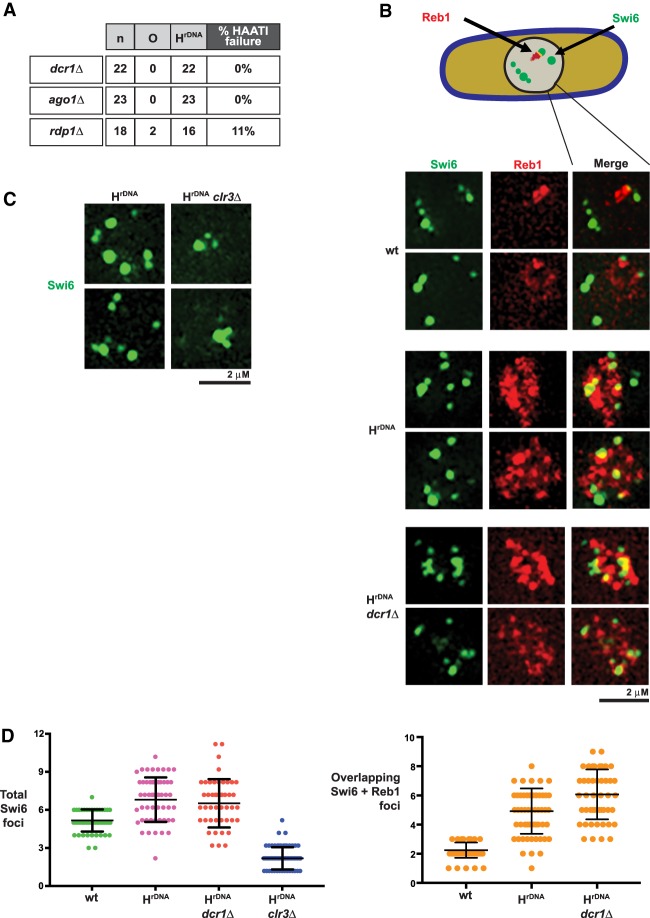
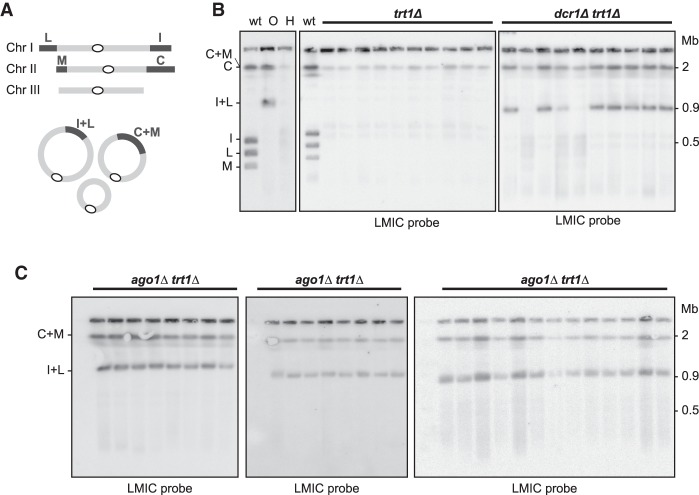
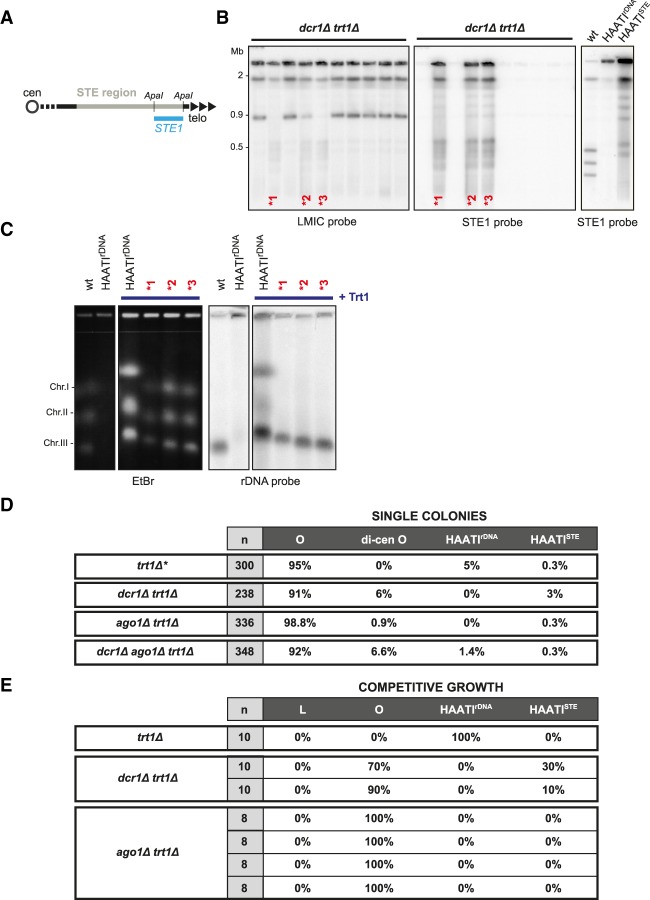
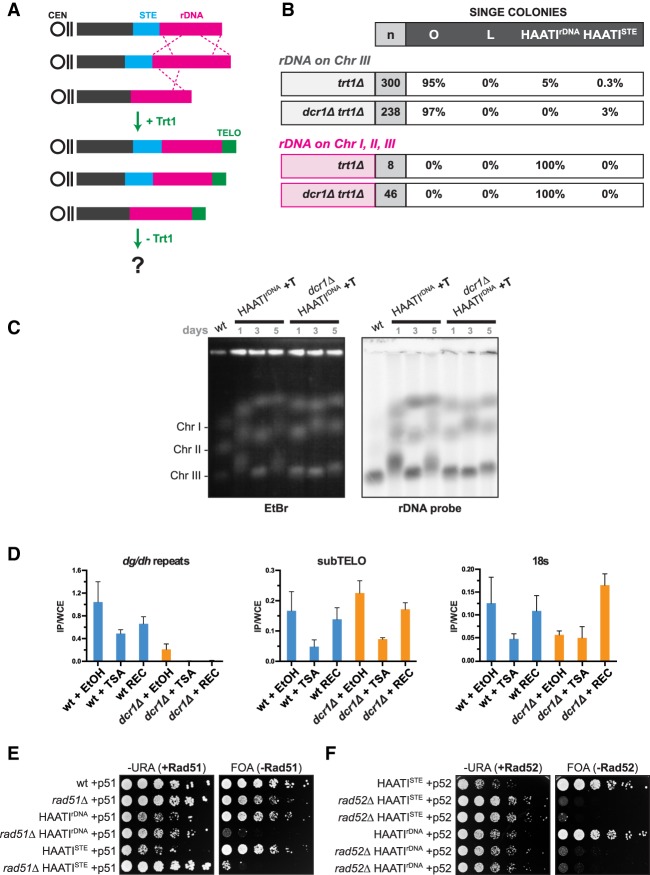
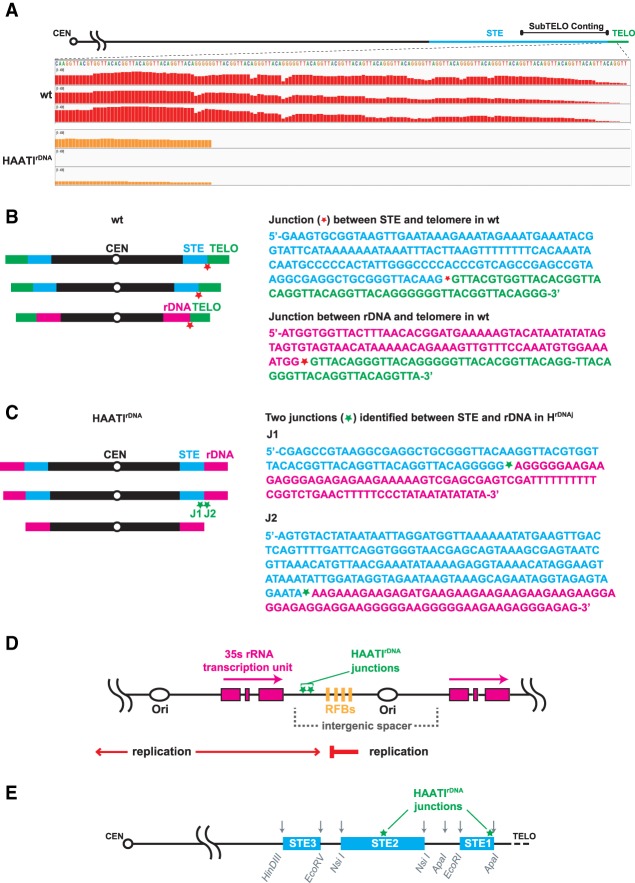
The identification of telomerase-negative HAATI (heterochromatin amplification-mediated and telomerase-independent) cells, in which telomeres are superseded by nontelomeric heterochromatin tracts, challenged the idea that canonical telomeres are essential for chromosome linearity and raised crucial questions as to how such tracts translocate to eroding chromosome ends and confer end protection. Here we show that HAATI arises when telomere loss triggers a newly recognized illegitimate translocation pathway that requires RNAi factors. While RNAi is necessary for the translocation events that mobilize ribosomal DNA (rDNA) tracts to all chromosome ends (forming "HAATI" chromosomes), it is dispensable for HAATI maintenance. Surprisingly, Dicer (Dcr1) plays a separate, RNAi-independent role in preventing formation of the rare HAATI subtype in which a different repetitive element (the subtelomeric element) replaces telomeres. Using genetics and fusions between shelterin components and rDNA-binding proteins, we mapped the mechanism by which rDNA loci engage crucial end protection factors-despite the absence of telomere repeats-and secure end protection. Sequence analysis of HAATI genomes allowed us to propose RNA and DNA polymerase template-switching models for the mechanism of RNAi-triggered rDNA translocations. Collectively, our results reveal unforeseen roles for noncoding RNAs (ncRNAs) in assembling a telomere-free chromosome end protection device.
端粒酶阴性 HAATI(异染色质扩增介导且端粒酶独立)细胞的鉴定,这些细胞的端粒被非端粒异染色质片段取代,这对经典端粒对于染色体线性至关重要的观点提出了挑战,并引发了一个关键问题,即这些片段如何转移到侵蚀性染色体末端并赋予末端保护。在这里,我们表明,当端粒丢失触发一种新识别的非法易位途径时,就会出现 HAATI,该途径需要 RNAi 因子。虽然 RNAi 对于将核糖体 DNA(rDNA)片段动员到所有染色体末端(形成“HAATI”染色体)的易位事件是必需的,但对于 HAATI 的维持却是可有可无的。令人惊讶的是,Dicer(Dcr1)在防止形成罕见的 HAATI 亚型中发挥着独立于 RNAi 的作用,在这种亚型中,不同的重复元件(端粒外元件)取代了端粒。通过遗传学和庇护素成分与 rDNA 结合蛋白之间的融合,我们确定了 rDNA 基因座与关键末端保护因子结合的机制,尽管缺乏端粒重复序列,但仍能确保末端保护。对 HAATI 基因组的序列分析使我们能够提出 RNA 和 DNA 聚合酶模板转换模型,用于解释 RNAi 触发的 rDNA 易位的机制。总的来说,我们的结果揭示了非编码 RNA(ncRNA)在组装无端粒染色体末端保护装置中的意想不到的作用。