Department of Cellular Biochemistry, Graduate School of Pharmaceutical Sciences, Kyushu University, 3-1-1 Maidashi, Higashiku, Fukuoka 812-8582, Japan.
Division of Transcriptomics, Medical Institute of Bioregulation, Kyushu University, 3-1-1 Maidashi, Higashiku, Fukuoka 812-8582, Japan.
Nucleic Acids Res. 2018 Jul 27;46(13):6683-6696. doi: 10.1093/nar/gky476.
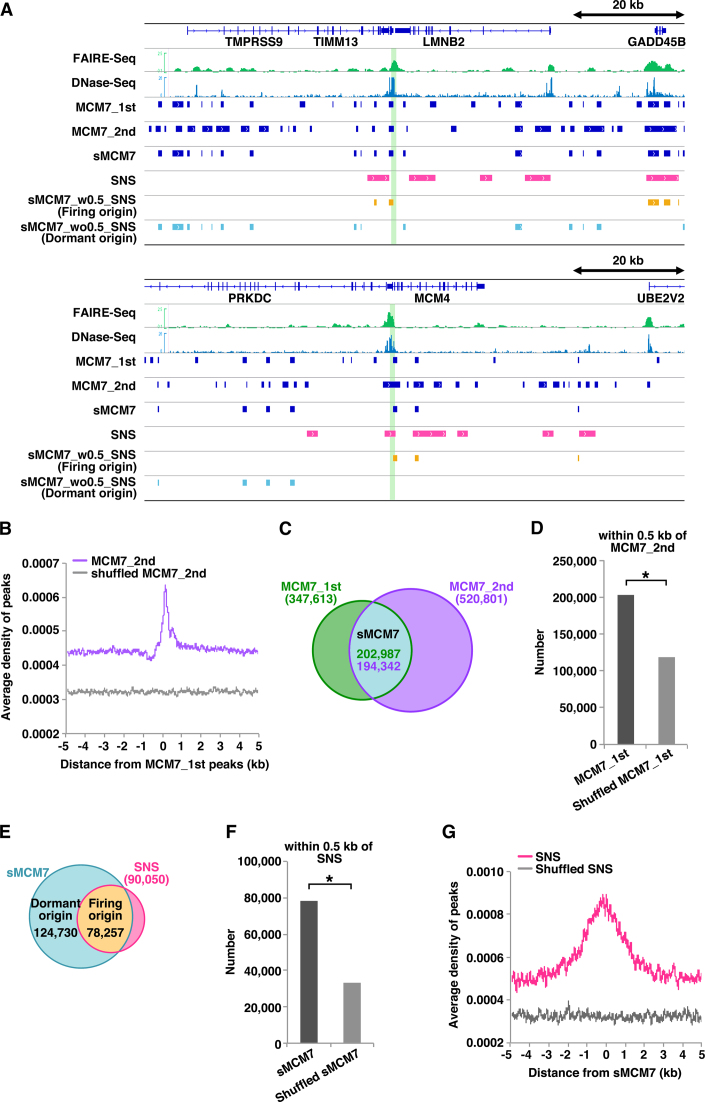
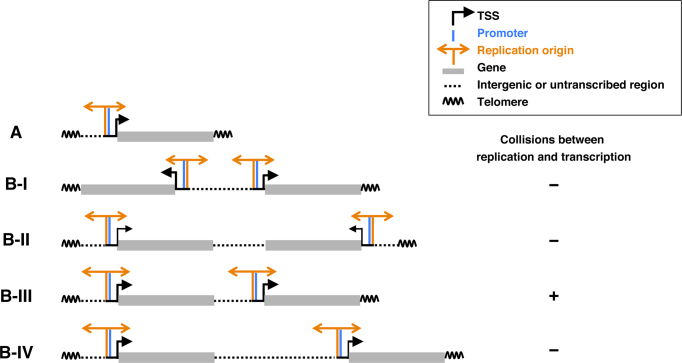
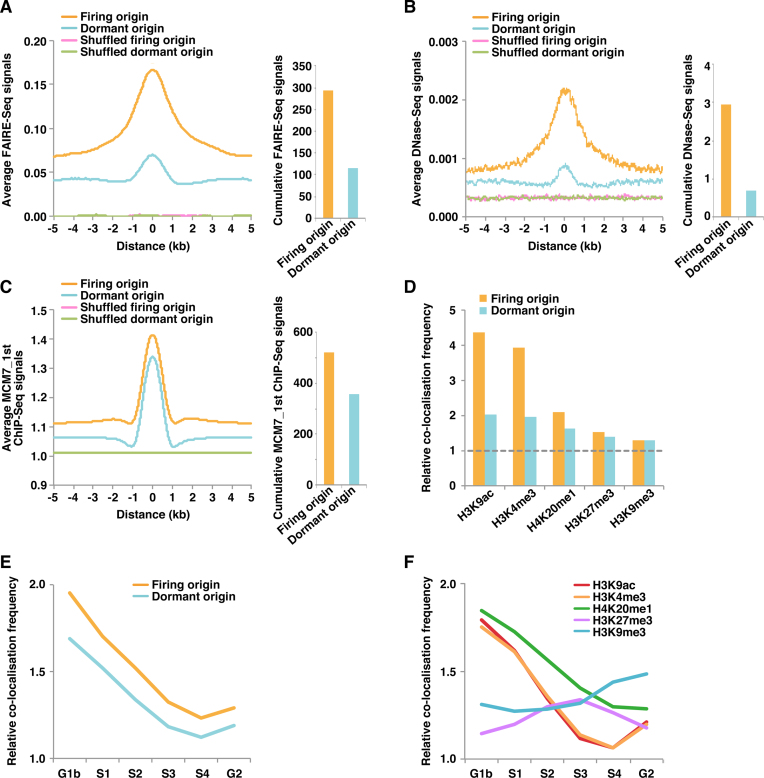
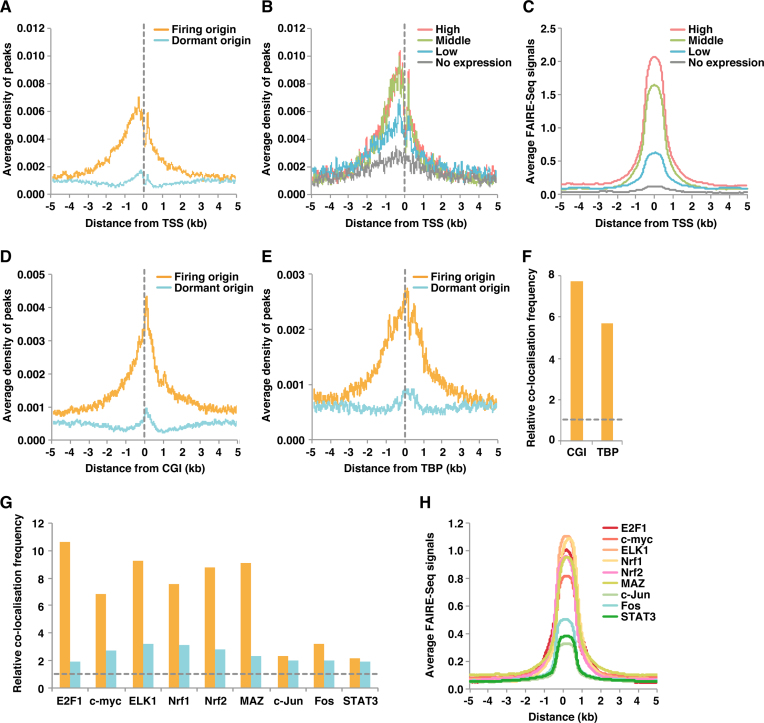
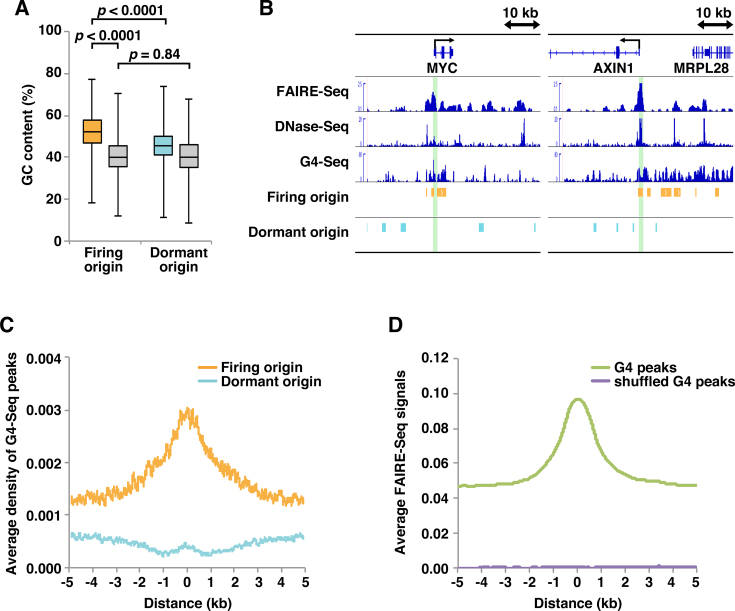
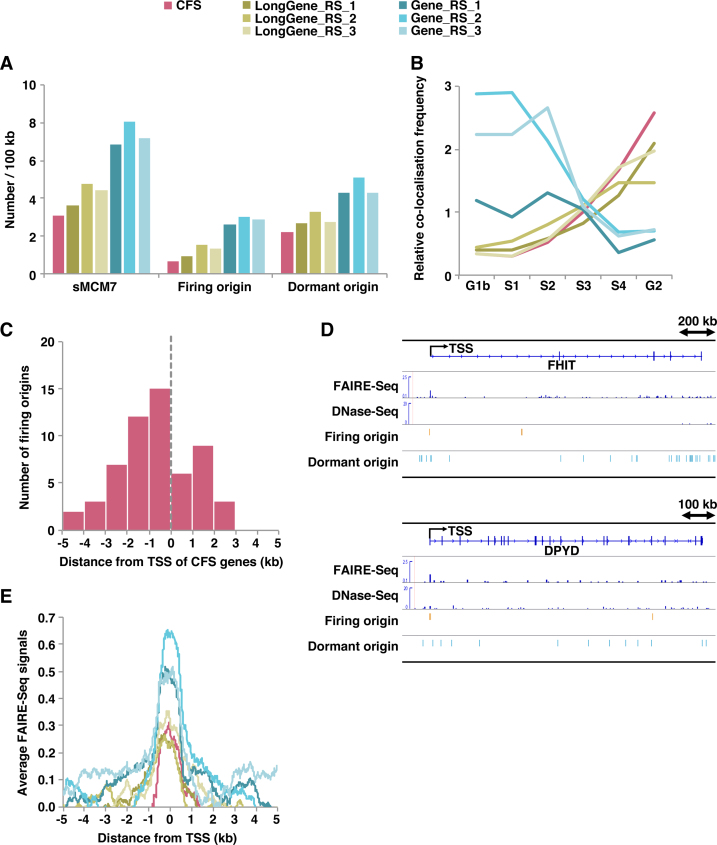
In metazoan cells, only a limited number of mini chromosome maintenance (MCM) complexes are fired during S phase, while the majority remain dormant. Several methods have been used to map replication origins, but such methods cannot identify dormant origins. Herein, we determined MCM7-binding sites in human cells using ChIP-Seq, classified them into firing and dormant origins using origin data and analysed their association with various chromatin signatures. Firing origins, but not dormant origins, were well correlated with open chromatin regions and were enriched upstream of transcription start sites (TSSs) of transcribed genes. Aggregation plots of MCM7 signals revealed minimal difference in the efficacy of MCM loading between firing and dormant origins. We also analysed common fragile sites (CFSs) and found a low density of origins at these sites. Nevertheless, firing origins were enriched upstream of the TSSs. Based on the results, we propose a model in which excessive MCMs are actively loaded in a genome-wide manner, irrespective of chromatin status, but only a fraction are passively fired in chromatin areas with an accessible open structure, such as regions upstream of TSSs of transcribed genes. This plasticity in the specification of replication origins may minimize collisions between replication and transcription.
在真核细胞中,只有有限数量的微小染色体维持(MCM)复合物在 S 期被激活,而大多数处于休眠状态。已经使用了几种方法来绘制复制原点,但这些方法无法识别休眠原点。在此,我们使用 ChIP-Seq 确定了人细胞中的 MCM7 结合位点,将它们分类为激活和休眠原点,并使用原点数据和分析它们与各种染色质特征的关联来分析它们。激活原点,但不是休眠原点,与开放染色质区域很好地相关,并且在转录起始位点(TSS)的转录基因上游富集。MCM7 信号的聚集图显示,激活和休眠原点之间的 MCM 加载效率几乎没有差异。我们还分析了常见的脆性位点(CFS),发现这些位点的原点密度较低。尽管如此,激活原点在 TSS 上游富集。基于这些结果,我们提出了一个模型,即无论染色质状态如何,过量的 MCM 都以全基因组的方式被主动加载,但只有一部分在具有可及开放结构的染色质区域中被被动激活,例如转录基因的 TSS 上游区域。这种复制原点的特异性可塑性可能最小化了复制和转录之间的碰撞。