Biomedical Sciences Research Complex, School of Biology, University of St Andrews, St Andrews, United Kingdom.
Elife. 2018 Jul 2;7:e36734. doi: 10.7554/eLife.36734.
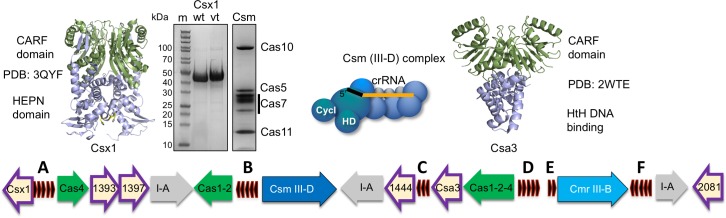
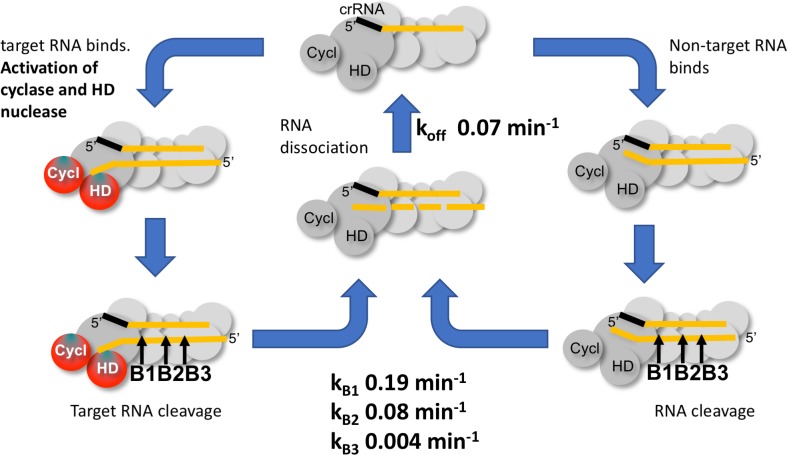
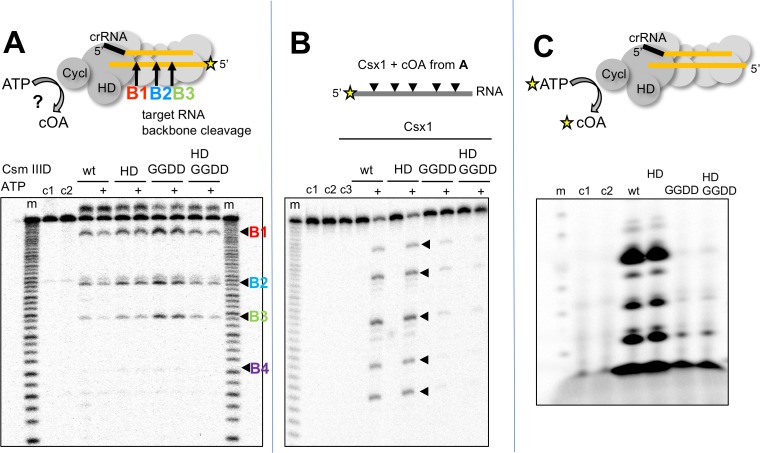
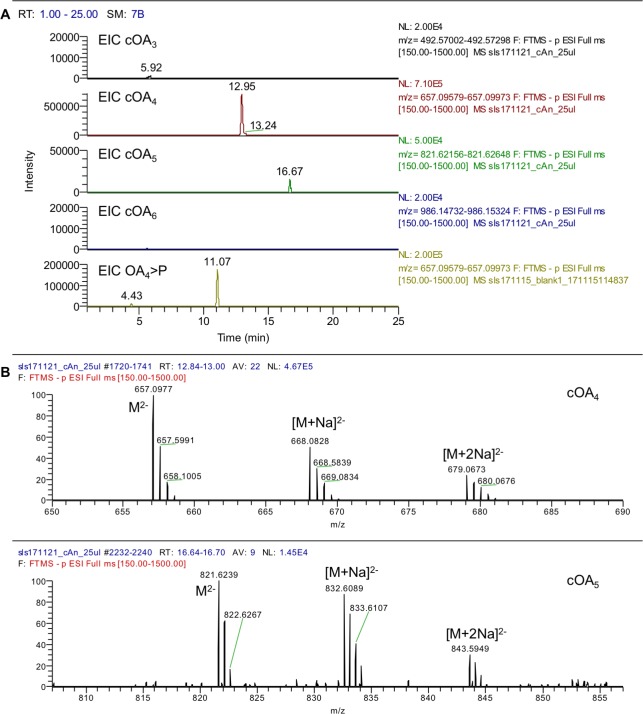
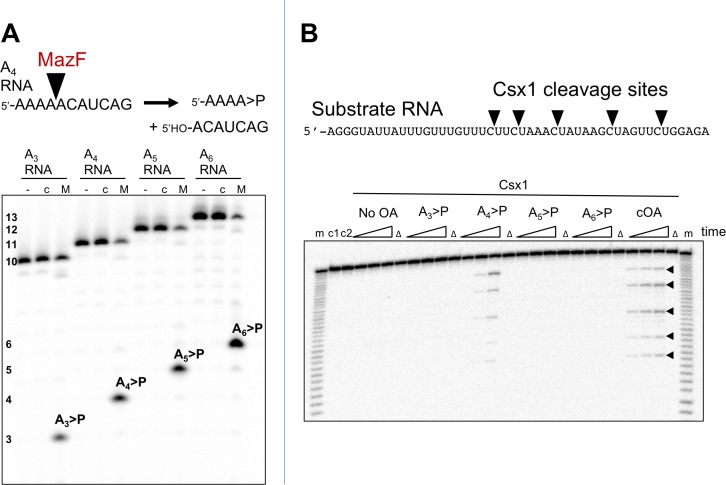
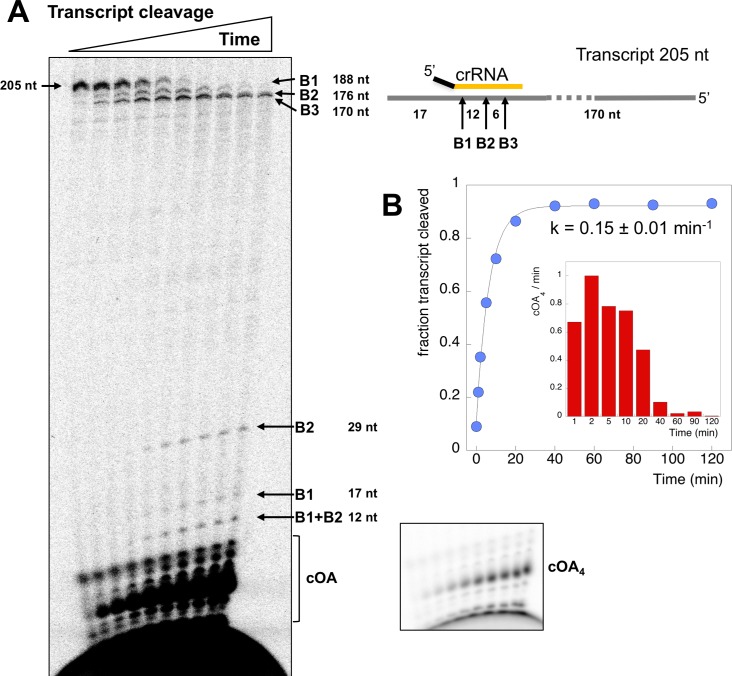
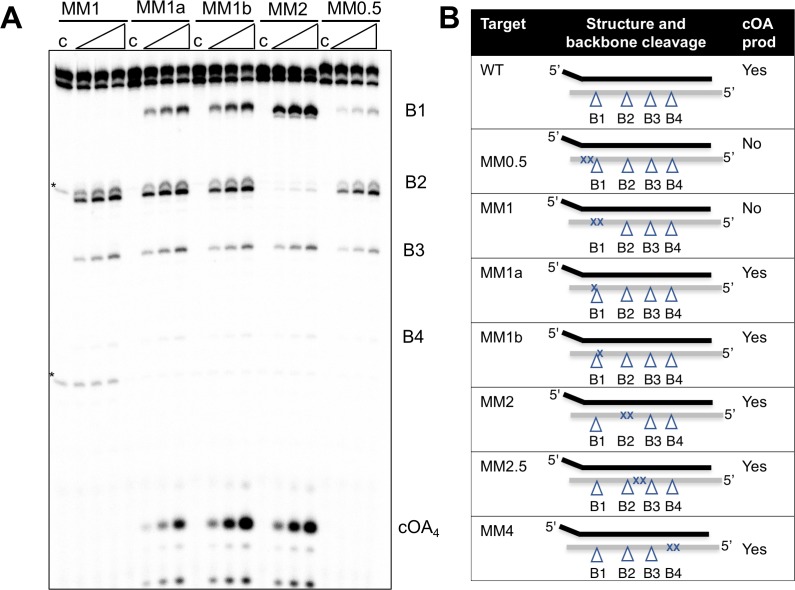
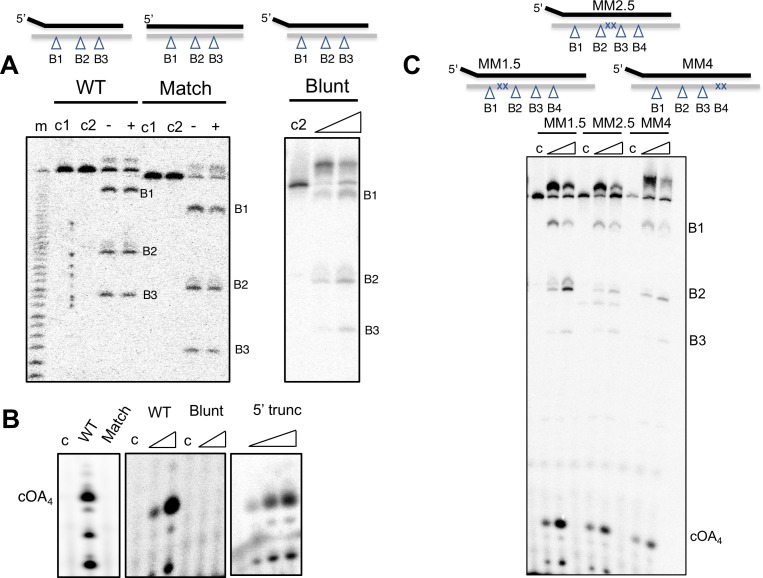
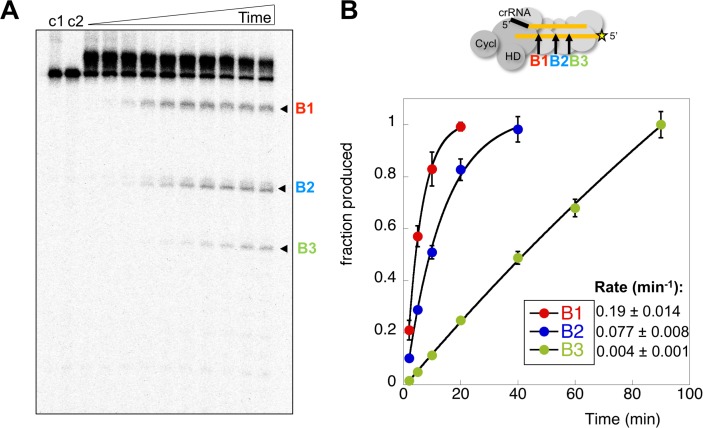
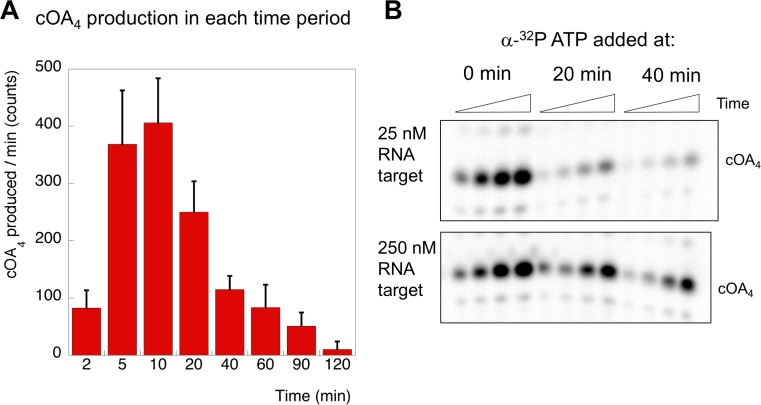
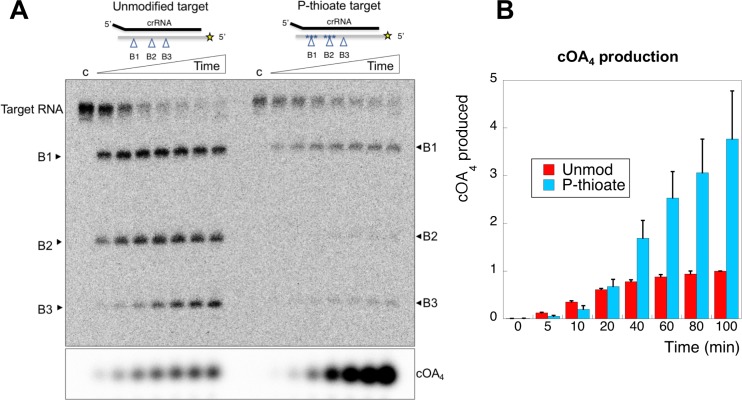
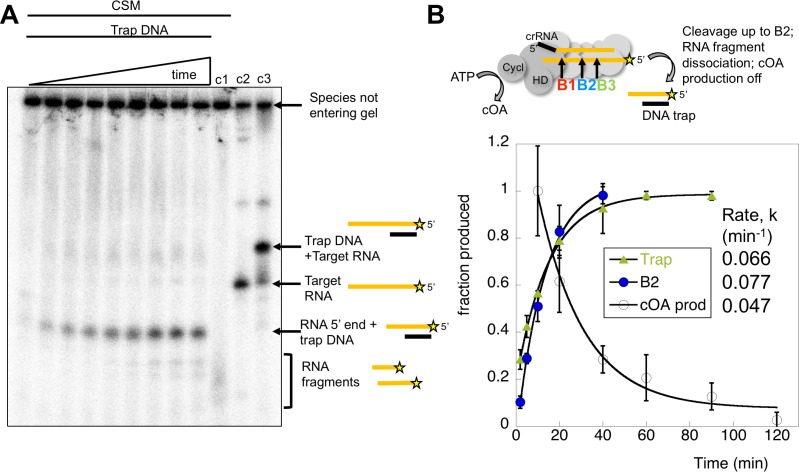
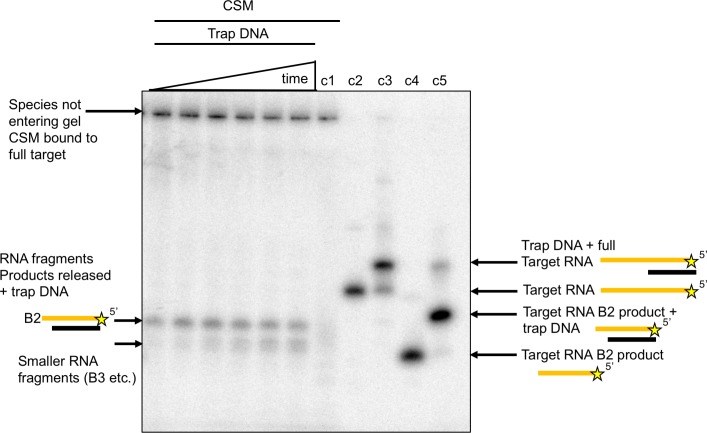
The CRISPR system for prokaryotic adaptive immunity provides RNA-mediated protection from viruses and mobile genetic elements. When viral RNA transcripts are detected, type III systems adopt an activated state that licenses DNA interference and synthesis of cyclic oligoadenylate (cOA). cOA activates nucleases and transcription factors that orchestrate the antiviral response. We demonstrate that cOA synthesis is subject to tight temporal control, commencing on target RNA binding, and is deactivated rapidly as target RNA is cleaved and dissociates. Mismatches in the target RNA are well tolerated and still activate the cyclase domain, except when located close to the 3' end of the target. Phosphorothioate modification reduces target RNA cleavage and stimulates cOA production. The 'RNA shredding' activity originally ascribed to type III systems may thus be a reflection of an exquisite mechanism for control of the Cas10 subunit, rather than a direct antiviral defence.
原核生物适应性免疫的 CRISPR 系统提供了 RNA 介导的抗病毒和移动遗传元件的保护。当检测到病毒 RNA 转录本时,III 型系统采用激活状态,许可 DNA 干扰和环寡腺苷酸(cOA)的合成。cOA 激活核酸酶和转录因子,协调抗病毒反应。我们证明 cOA 合成受到严格的时间控制,起始于靶 RNA 结合,并随着靶 RNA 的切割和解离迅速失活。靶 RNA 中的错配得到很好的容忍,并且仍然激活环化酶结构域,除非位于靶 RNA 的 3' 端附近。硫代磷酸酯修饰降低了靶 RNA 的切割,并刺激 cOA 的产生。因此,最初归因于 III 型系统的“RNA 粉碎”活性可能反映了对 Cas10 亚基的精细控制机制,而不是直接的抗病毒防御。