Molecular and Cellular Biology Graduate Program, University of Massachusetts, Amherst, United States.
Department of Biological Sciences, Dartmouth College, Hanover, United States.
Elife. 2018 Aug 7;7:e36745. doi: 10.7554/eLife.36745.
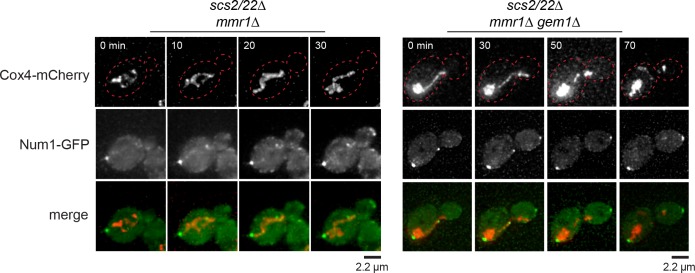
Cortical dynein generates pulling forces via microtubule (MT) end capture-shrinkage and lateral MT sliding mechanisms. In , the dynein attachment molecule Num1 interacts with endoplasmic reticulum (ER) and mitochondria to facilitate spindle positioning across the mother-bud neck, but direct evidence for how these cortical contacts regulate dynein-dependent pulling forces is lacking. We show that loss of Scs2/Scs22, ER tethering proteins, resulted in defective Num1 distribution and loss of dynein-dependent MT sliding, the hallmark of dynein function. Cells lacking Scs2/Scs22 performed spindle positioning via MT end capture-shrinkage mechanism, requiring dynein anchorage to an ER- and mitochondria-independent population of Num1, dynein motor activity, and CAP-Gly domain of dynactin Nip100/p150 subunit. Additionally, a CAAX-targeted Num1 rescued loss of lateral patches and MT sliding in the absence of Scs2/Scs22. These results reveal distinct populations of Num1 and underline the importance of their spatial distribution as a critical factor for regulating dynein pulling force.
皮层动力蛋白通过微管(MT)末端捕获-收缩和横向 MT 滑动机制产生拉力。在 ,动力蛋白附着分子 Num1 与内质网(ER)和线粒体相互作用,以促进纺锤体在母-芽颈部的定位,但缺乏关于这些皮质接触如何调节依赖动力蛋白的拉力的直接证据。我们表明,ER 固定蛋白 Scs2/Scs22 的缺失导致 Num1 分布缺陷和依赖动力蛋白的 MT 滑动丧失,这是动力蛋白功能的标志。缺乏 Scs2/Scs22 的细胞通过 MT 末端捕获-收缩机制进行纺锤体定位,需要动力蛋白锚定在 ER 和线粒体独立的 Num1 群体、动力蛋白马达活性以及 dynactin Nip100/p150 亚基的 CAP-Gly 结构域。此外,靶向 CAAX 的 Num1 挽救了 Scs2/Scs22 缺失时横向斑块和 MT 滑动的缺失。这些结果揭示了 Num1 的不同群体,并强调了它们的空间分布作为调节动力蛋白拉力的关键因素的重要性。