Biotechnology and Nuclear Technology Research Institute, Sichuan Academy of Agricultural Sciences, Chengdu, 610061, China.
Center of Analysis and Testing, Sichuan Academy of Agricultural Sciences, Chengdu, 610061, China.
BMC Plant Biol. 2018 Sep 5;18(1):181. doi: 10.1186/s12870-018-1399-x.
Sweetpotato (Ipomoea batatas (L.) Lam.) is one of the most important crops from the family of Convolvulaceae. It is widely reported that cultivated sweetpotato was originated from Ipomoea trifida. However, diploid, tetraploid and hexaploid I. trifida were found in nature. The relationship, between them, and among them and sweetpotato, is remaining unclear.
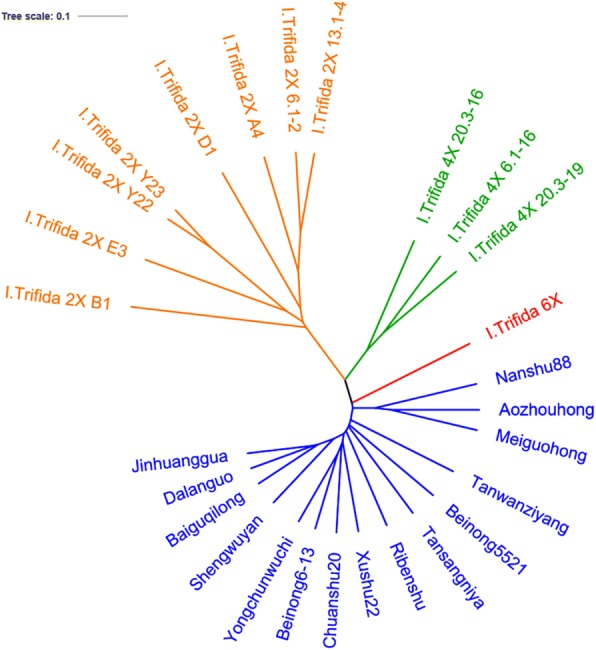
In the present study, we detected the genome diversity and relationship of sweetpotato and different polyploidy types I. trifida using Restriction-site Associated DNA Sequencing (RAD-seq). A total of 38,605 RAD-tags containing 832,204 SNPs had been identified. These tags were annotated using five public databases, about 11,519 tags were aligned to functional genes in various pathways. Based on SNP genotype, phylogenetic relation analysis results confirmed that cultivated sweetpotato has a closer relationship with I. trifida 6× than with I. trifida 4X and I. trifida 2×. Besides, 5042 SSRs were detected in I. trifida 6×, and 3202 pairs of high-quality SSR primers were developed. A total of 68 primers were randomly selected and synthesized, of which 61 were successfully amplified.
These results provided new evidence that cultivated sweetpotato originated from I. trifida 6×, and that I. trifida 6× evolved from I. trifida 4X and I. trifida 2×. Therefore, using I. trifida 6× as the model plant of sweetpotato research should be more practical than using I. trifida 2× in the future. Meanwhile, sequence information and markers from the present study will be helpful for sweetpotato and I. trifida studies in the future.
番薯(Ipomoea batatas (L.) Lam.)是旋花科中最重要的作物之一。据广泛报道,栽培番薯起源于Ipomoea trifida。然而,自然界中存在二倍体、四倍体和六倍体的 I. trifida。它们之间以及与番薯之间的关系尚不清楚。
在本研究中,我们使用限制性位点相关 DNA 测序(RAD-seq)检测了番薯和不同多倍体类型 I. trifida 的基因组多样性和关系。共检测到 38605 个包含 832204 个 SNP 的 RAD 标签。这些标签使用五个公共数据库进行注释,约 11519 个标签被映射到各种途径的功能基因上。基于 SNP 基因型,系统发育关系分析结果证实,栽培番薯与 6× I. trifida 的关系比与 4× I. trifida 和 2× I. trifida 的关系更为密切。此外,在 6× I. trifida 中检测到 5042 个 SSRs,并开发了 3202 对高质量 SSR 引物。随机选择并合成了 68 对引物,其中 61 对成功扩增。
这些结果提供了新的证据,表明栽培番薯起源于 6× I. trifida,而 6× I. trifida 是由 4× I. trifida 和 2× I. trifida 进化而来。因此,未来使用 6× I. trifida 作为番薯研究的模式植物比使用 2× I. trifida 更为实际。同时,本研究的序列信息和标记将有助于未来对番薯和 I. trifida 的研究。