Kremling Andreas, Geiselmann Johannes, Ropers Delphine, de Jong Hidde
Systems Biotechnology, Technical University of Munich, Boltzmannstrasse 15, Garching b. München, 85748, Germany.
Laboratoire Interdisciplinaire de Physique, Université Grenoble Alpes, 140 avenue de la Physique, Saint Martin d'Hères, 38402, France.
BMC Syst Biol. 2018 Sep 21;12(1):82. doi: 10.1186/s12918-018-0604-8.
Carbon catabolite repression (CCR) controls the order in which different carbon sources are metabolised. Although this system is one of the paradigms of regulation in bacteria, the underlying mechanisms remain controversial. CCR involves the coordination of different subsystems of the cell - responsible for the uptake of carbon sources, their breakdown for the production of energy and precursors, and the conversion of the latter to biomass. The complexity of this integrated system, with regulatory mechanisms cutting across metabolism, gene expression, and signalling, has motivated important modelling efforts over the past four decades, especially in the enterobacterium Escherichia coli.
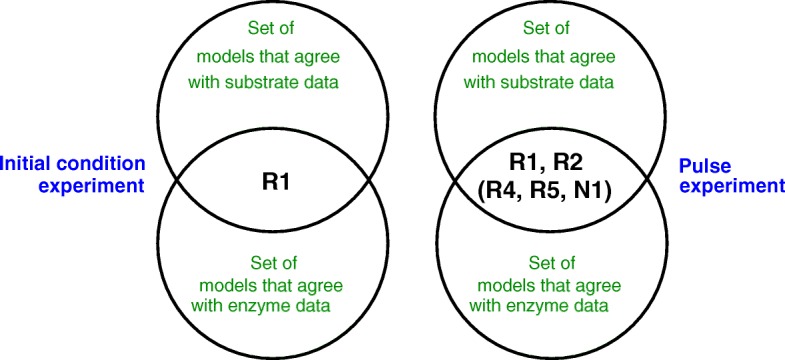
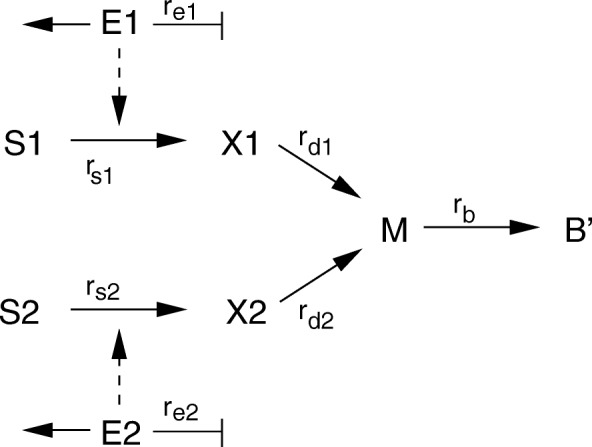
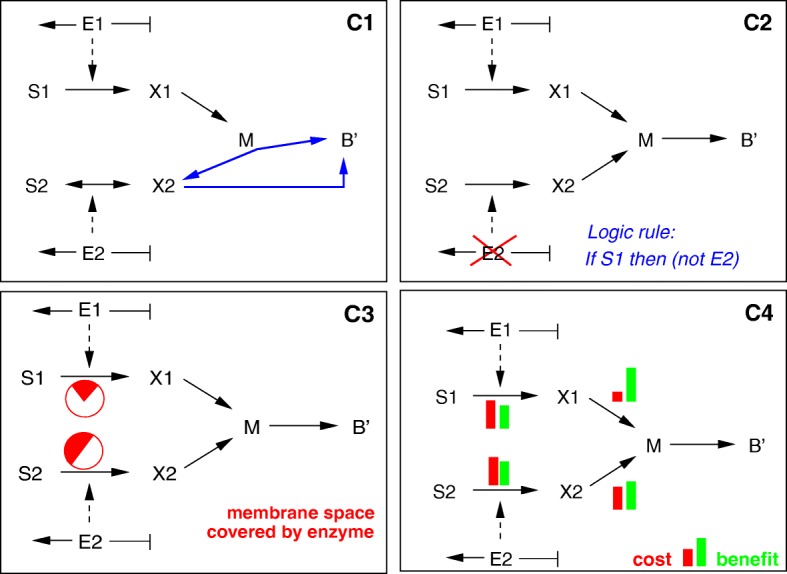
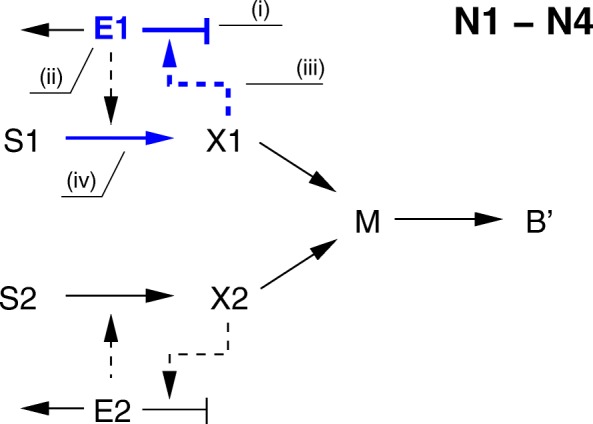
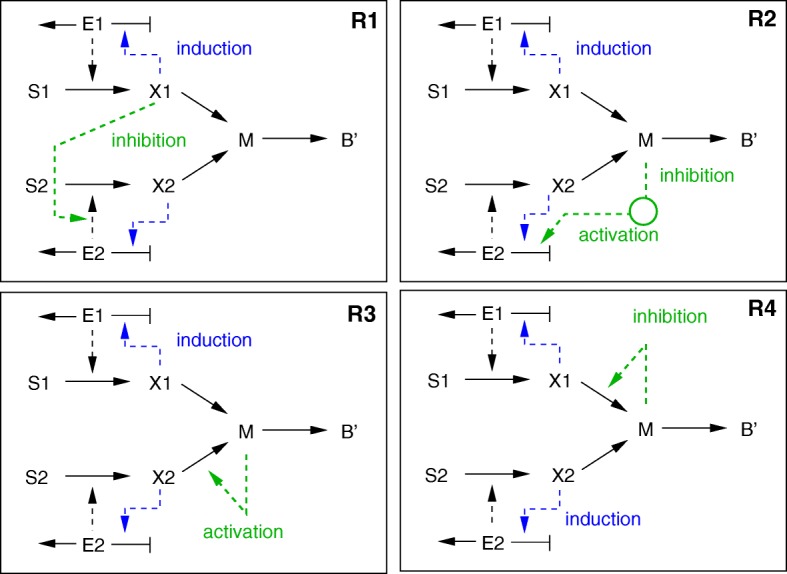
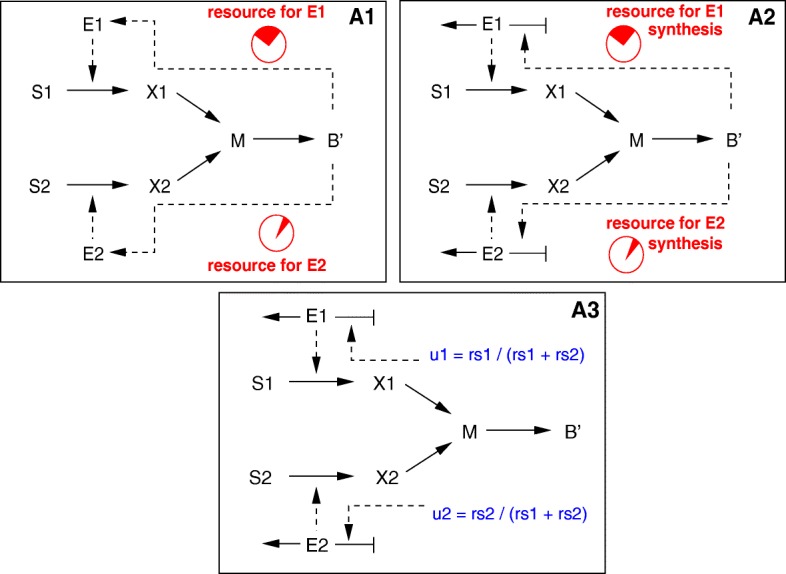
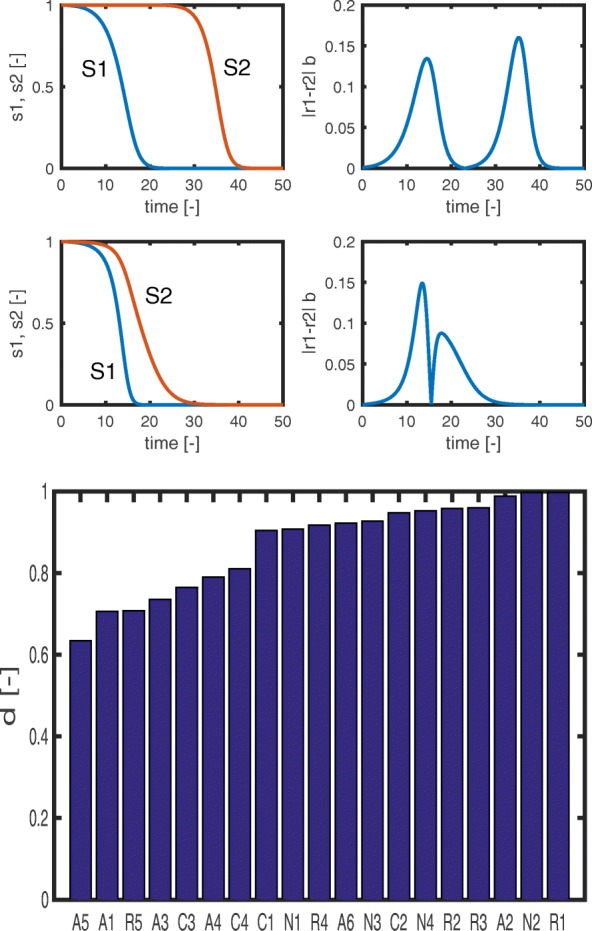
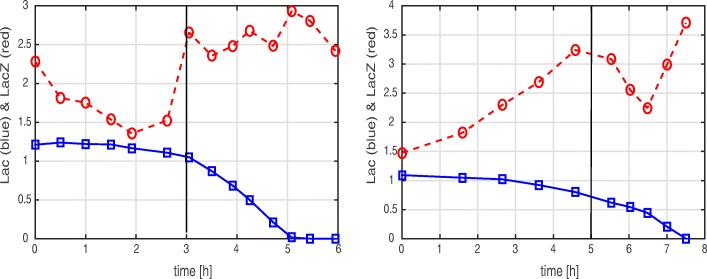
Starting from a simple core model with only four intracellular metabolites, we develop an ensemble of model variants, all showing diauxic growth behaviour during a batch process. The model variants fall into one of the four categories: flux balance models, kinetic models with growth dilution, kinetic models with regulation, and resource allocation models. The model variants differ from one another in only a single aspect, each breaking the symmetry between the two substrate assimilation pathways in a different manner, and can be quantitatively compared using a so-called diauxic growth index. For each of the model variants, we predict the behaviour in two new experimental conditions, namely a glucose pulse for a culture growing in minimal medium with lactose and a batch culture with different initial concentrations of the components of the transport systems. When qualitatively comparing these predictions with experimental data for these two conditions, a number of models can be excluded while other model variants are still not discriminable. The best-performing model variants are based on inducer inclusion and activation of enzymatic genes by a global transcription factor, but the other proposed factors may complement these well-known regulatory mechanisms.
The model ensemble presented here offers a better understanding of the variety of mechanisms that have been proposed to play a role in CCR. In addition, it provides an educational resource for systems biology, as it gives an introduction to a broad range of modeling approaches in the context of a simple but biologically relevant example.
碳分解代谢物阻遏(CCR)控制着不同碳源的代谢顺序。尽管该系统是细菌调控的范例之一,但其潜在机制仍存在争议。CCR涉及细胞不同子系统的协调——负责碳源的摄取、将其分解以产生能量和前体物质,以及将后者转化为生物量。这个整合系统的复杂性,其调控机制贯穿代谢、基因表达和信号传导,在过去四十年中推动了重要的建模工作,尤其是在肠道细菌大肠杆菌中。
从仅包含四种细胞内代谢物的简单核心模型出发,我们开发了一系列模型变体,所有这些变体在分批培养过程中均表现出双相生长行为。这些模型变体分为四类之一:通量平衡模型、具有生长稀释的动力学模型、具有调控的动力学模型和资源分配模型。这些模型变体仅在一个方面彼此不同,每个变体以不同方式打破了两种底物同化途径之间的对称性,并且可以使用所谓的双相生长指数进行定量比较。对于每个模型变体,我们预测其在两种新实验条件下的行为,即对于在含有乳糖的基本培养基中生长的培养物的葡萄糖脉冲,以及具有不同初始浓度转运系统组分的分批培养。当将这些预测与这两种条件下的实验数据进行定性比较时,可以排除一些模型,而其他模型变体仍然无法区分。表现最佳的模型变体基于诱导物的纳入以及全局转录因子对酶基因的激活,但其他提出的因素可能补充这些众所周知的调控机制。
这里提出的模型集合有助于更好地理解已被提出在CCR中起作用的各种机制。此外,它为系统生物学提供了一种教育资源,因为它在一个简单但与生物学相关的例子的背景下介绍了广泛的建模方法。