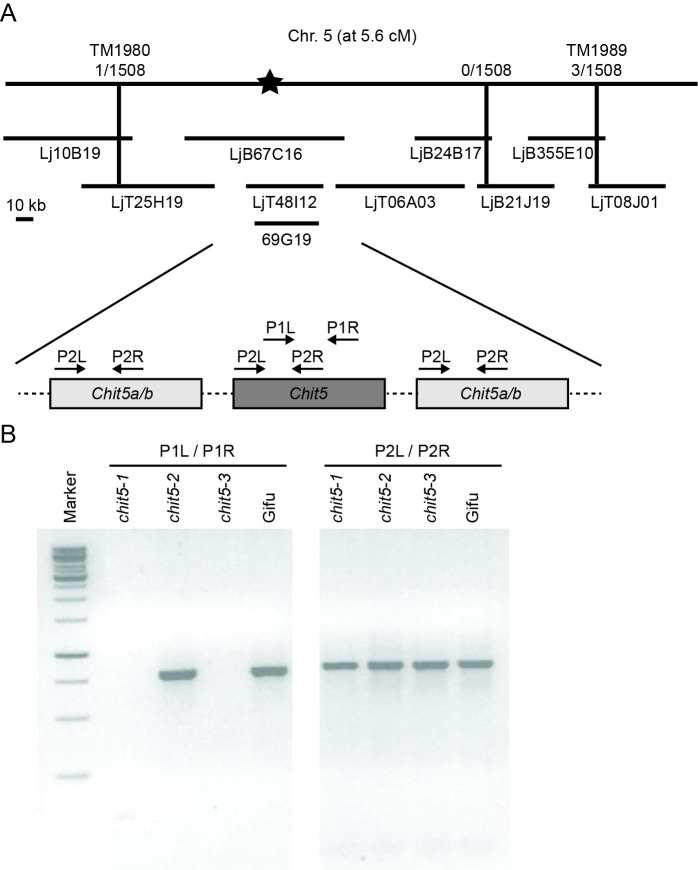
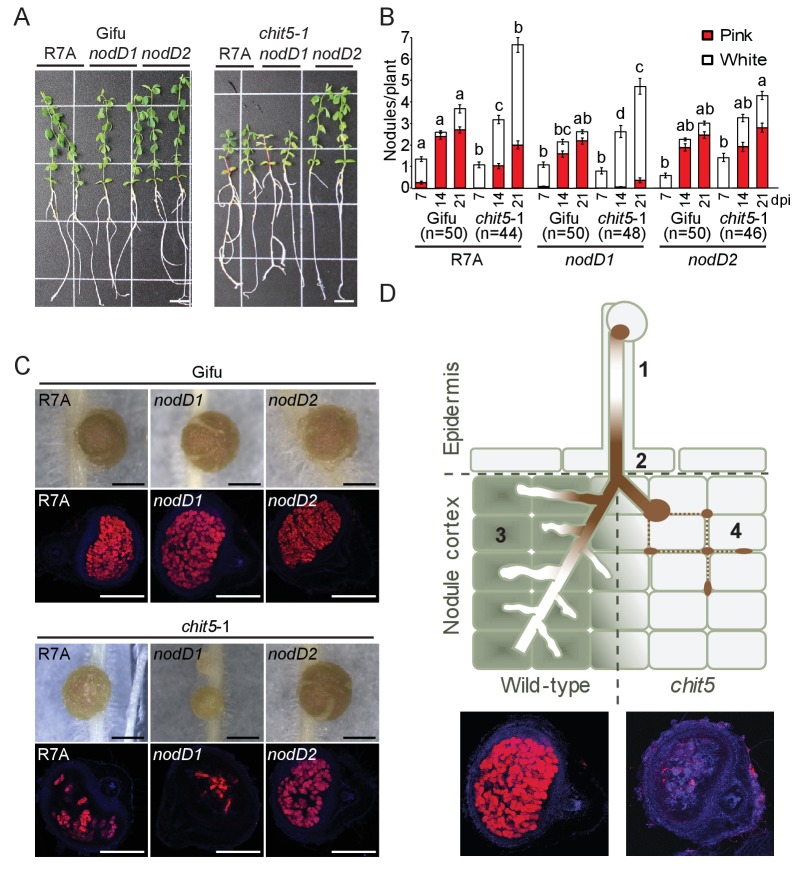
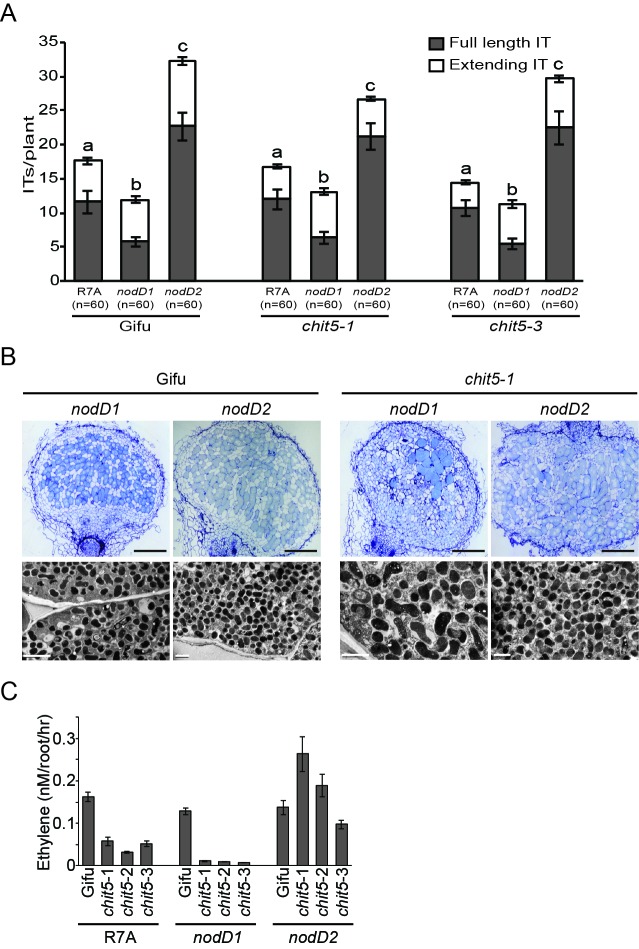
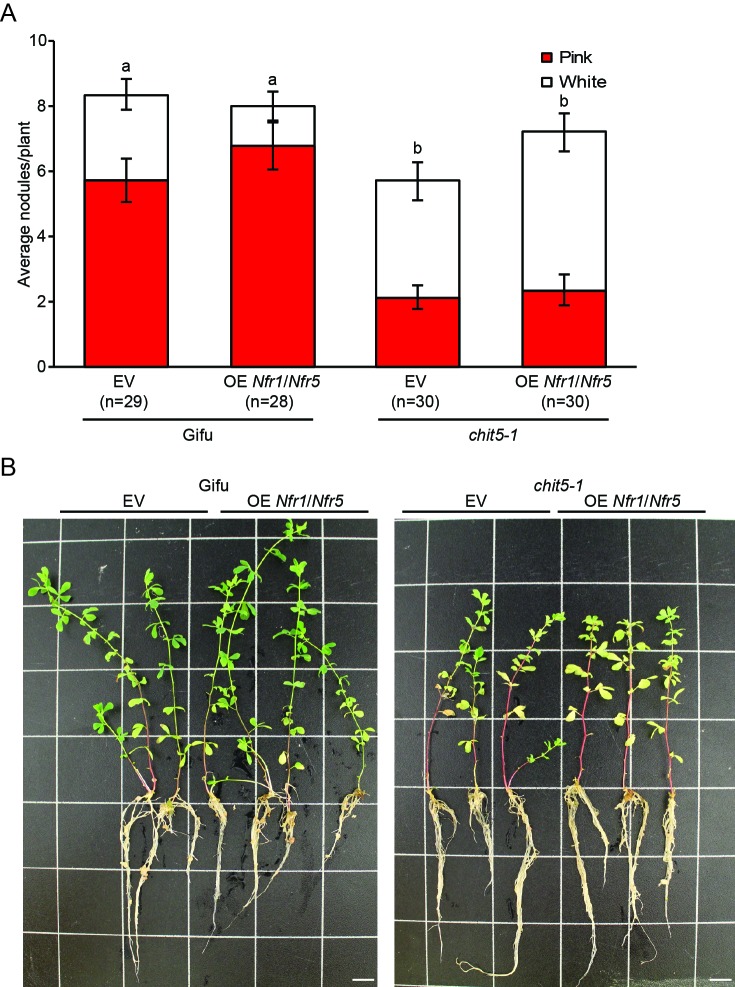
Department of Molecular Biology and Genetics, Aarhus University, Aarhus, Denmark.
Department of Chemistry, University of Copenhagen, Frederiksberg, Denmark.
Elife. 2018 Oct 4;7:e38874. doi: 10.7554/eLife.38874.
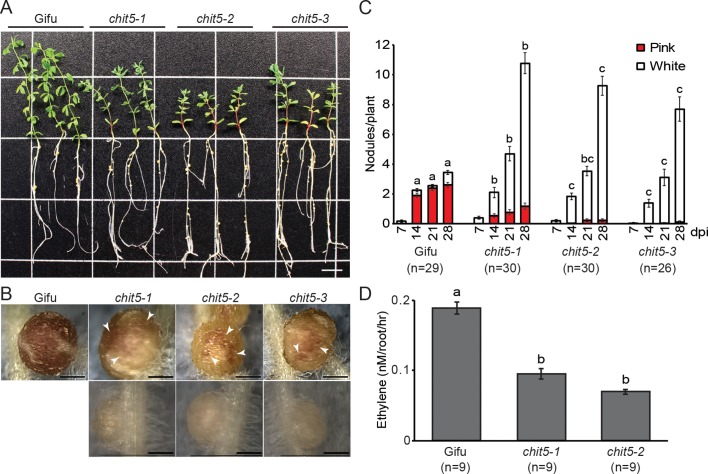
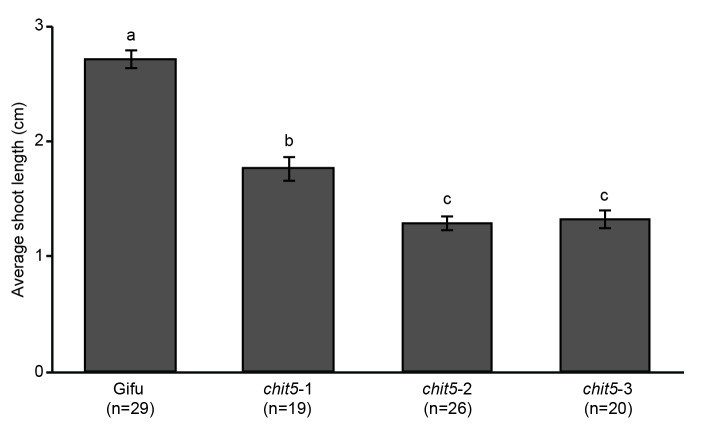
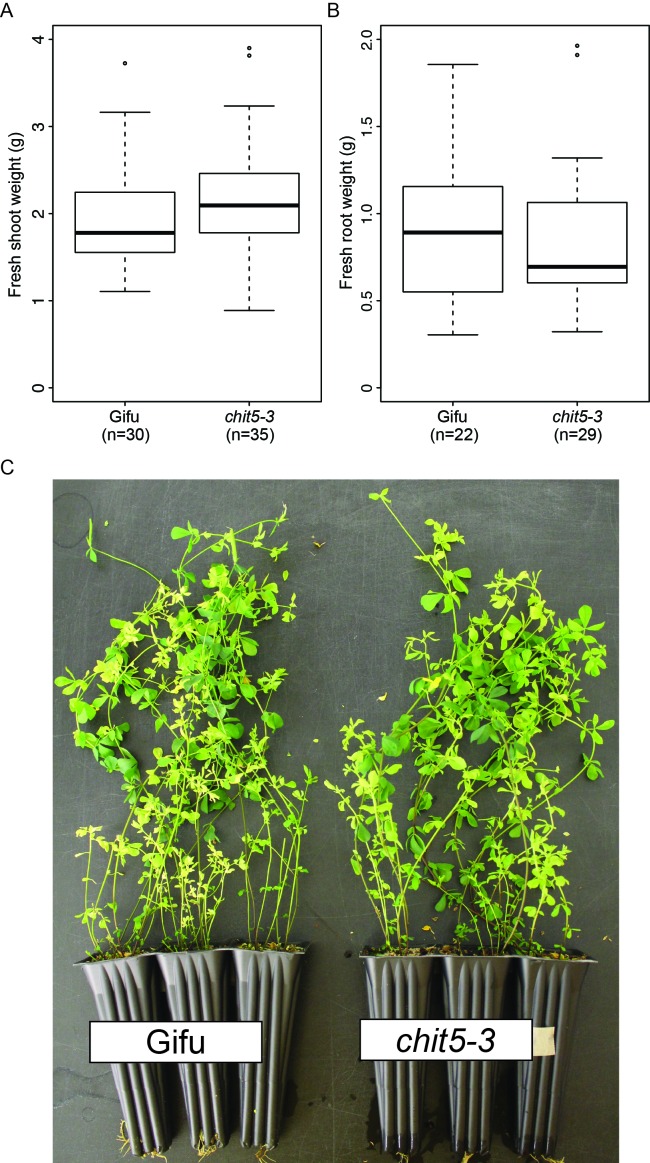
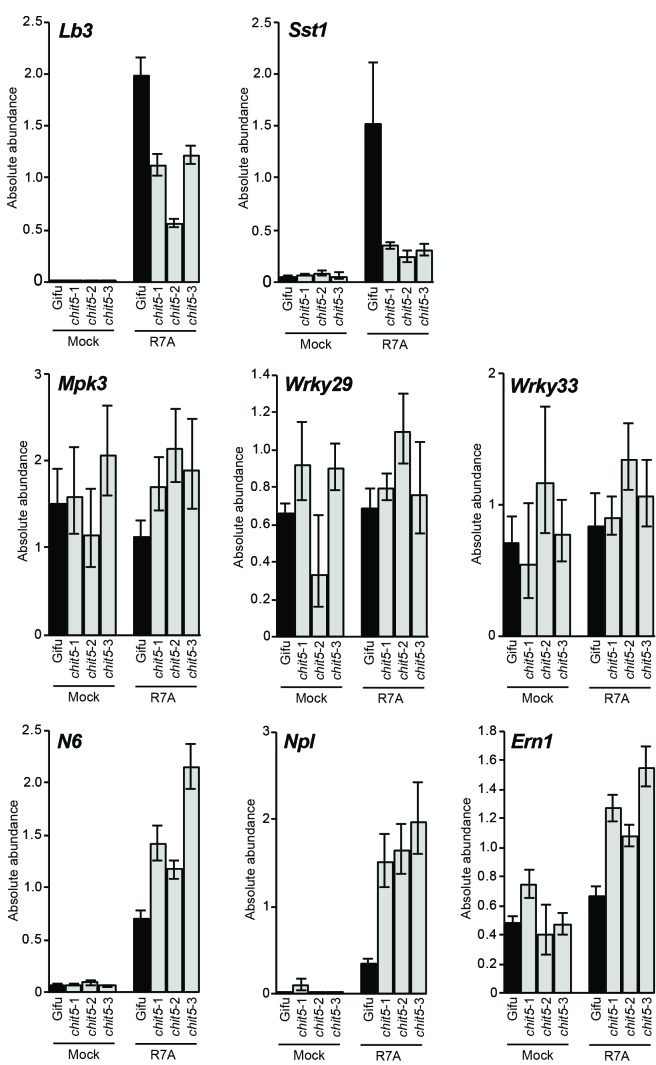
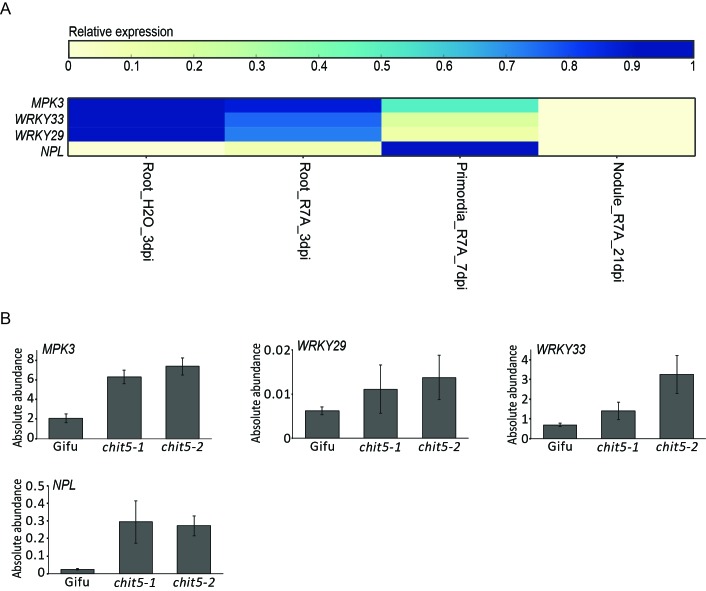
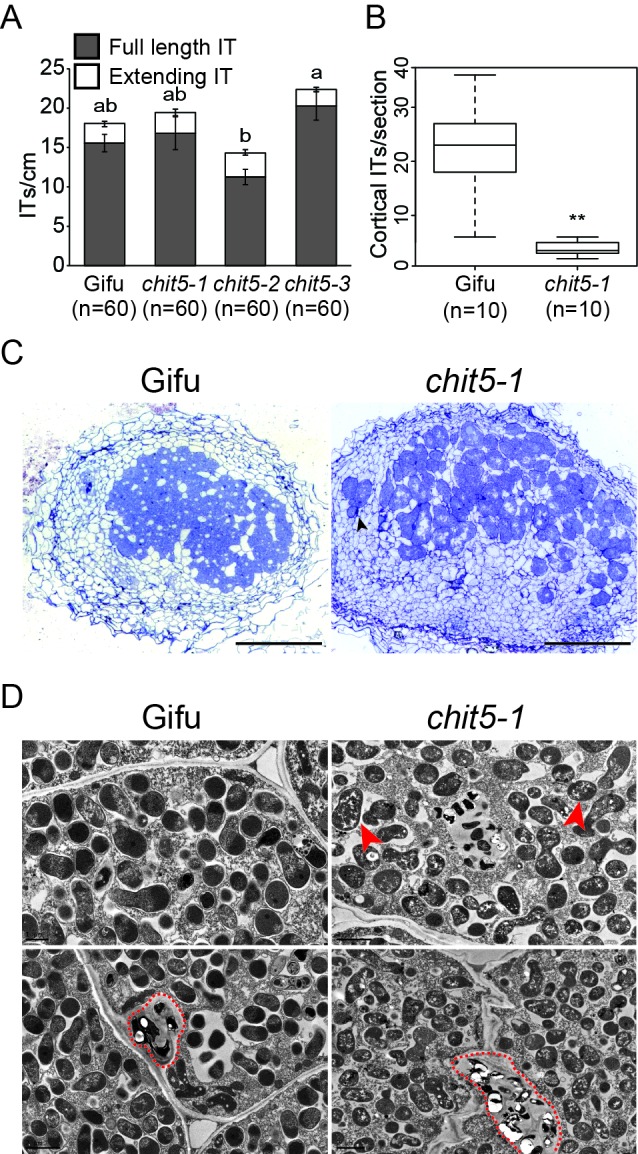
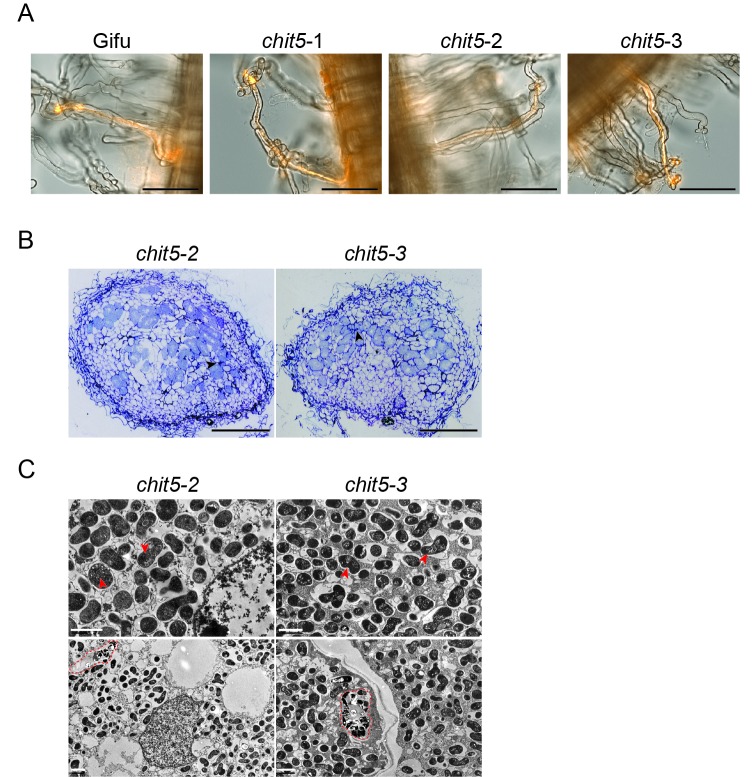
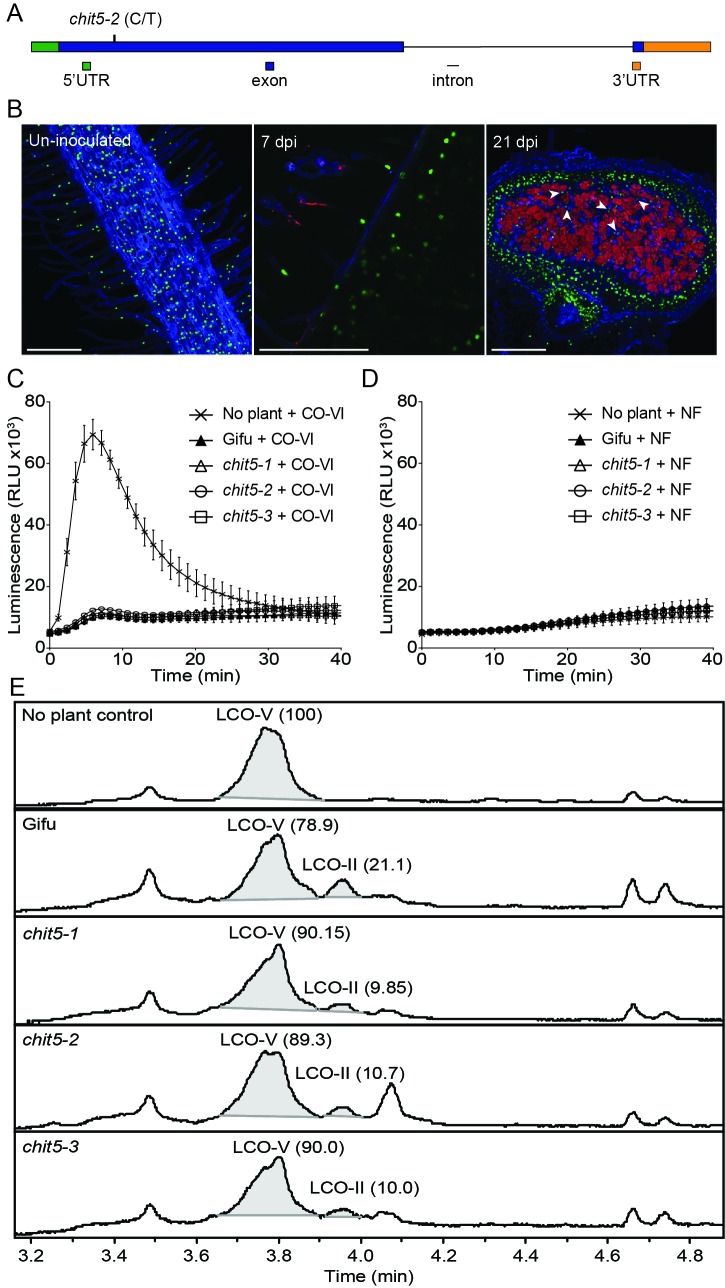
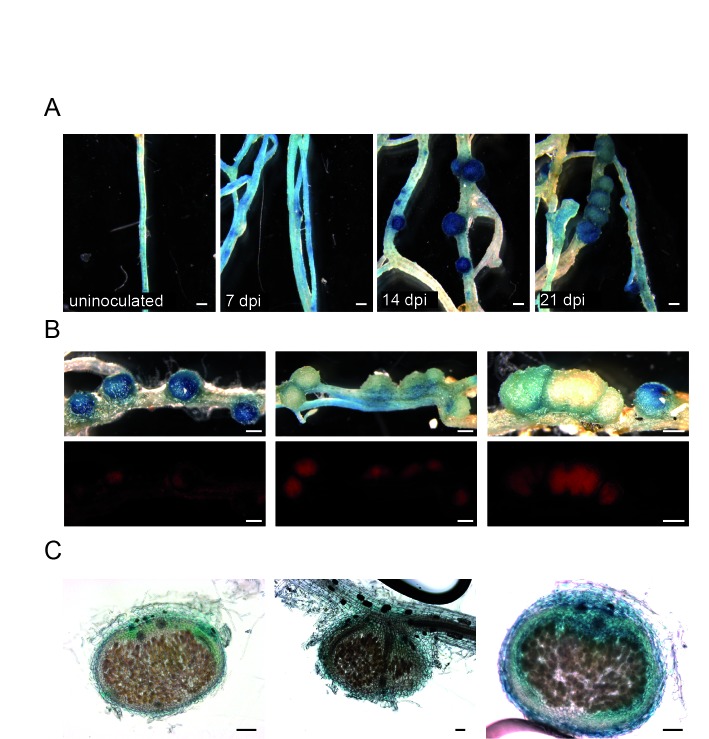
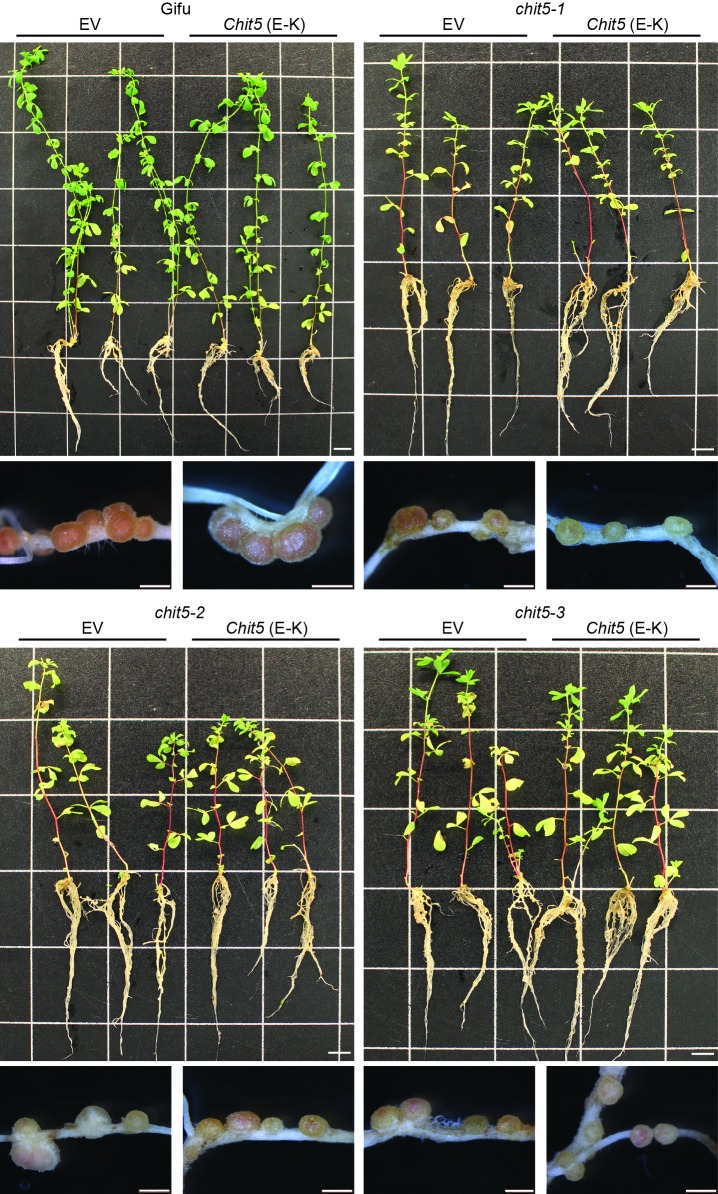
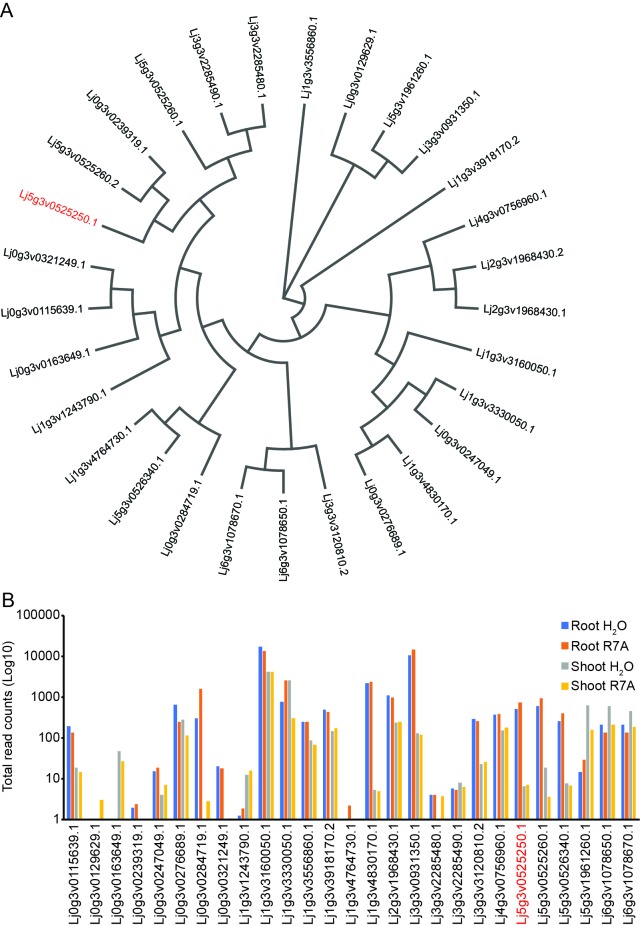
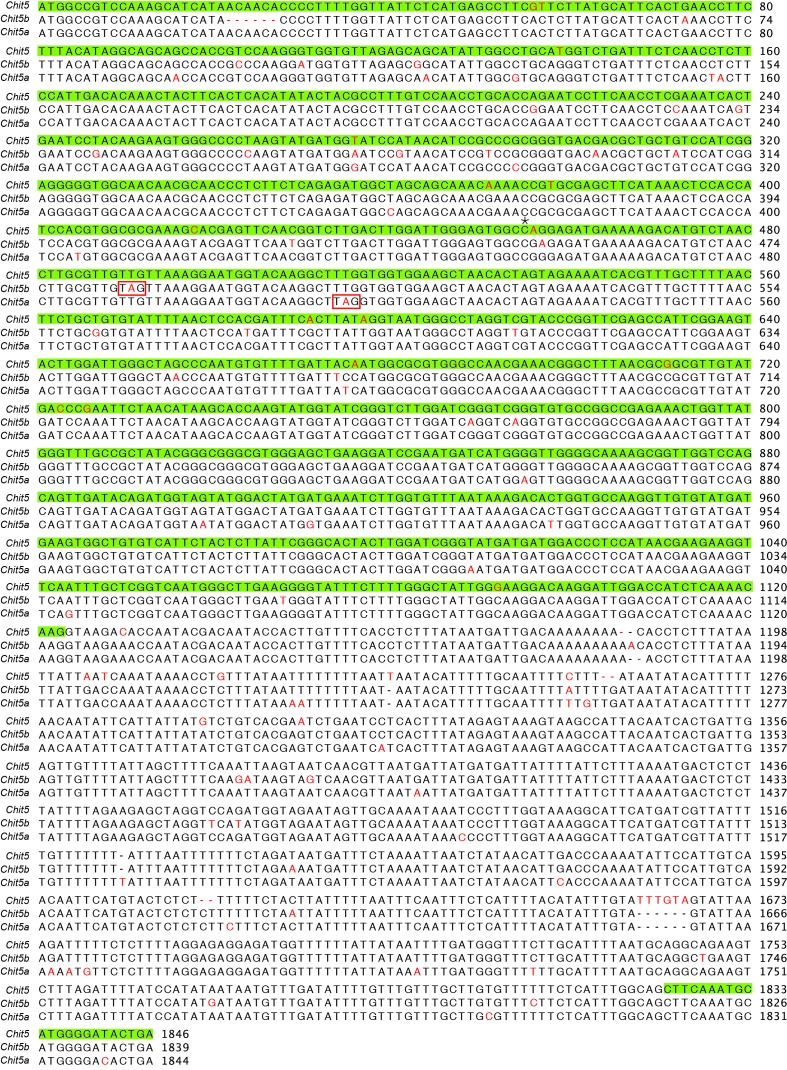
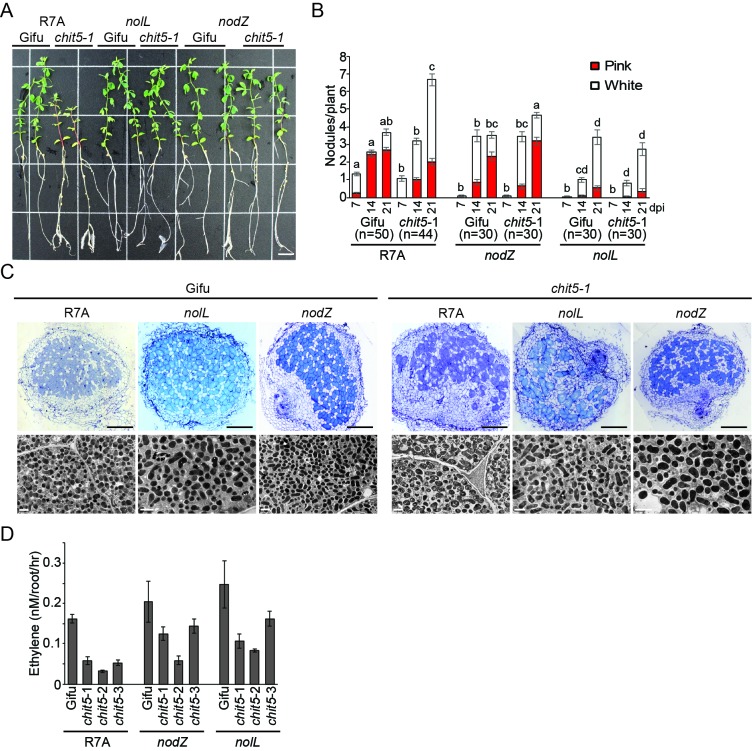
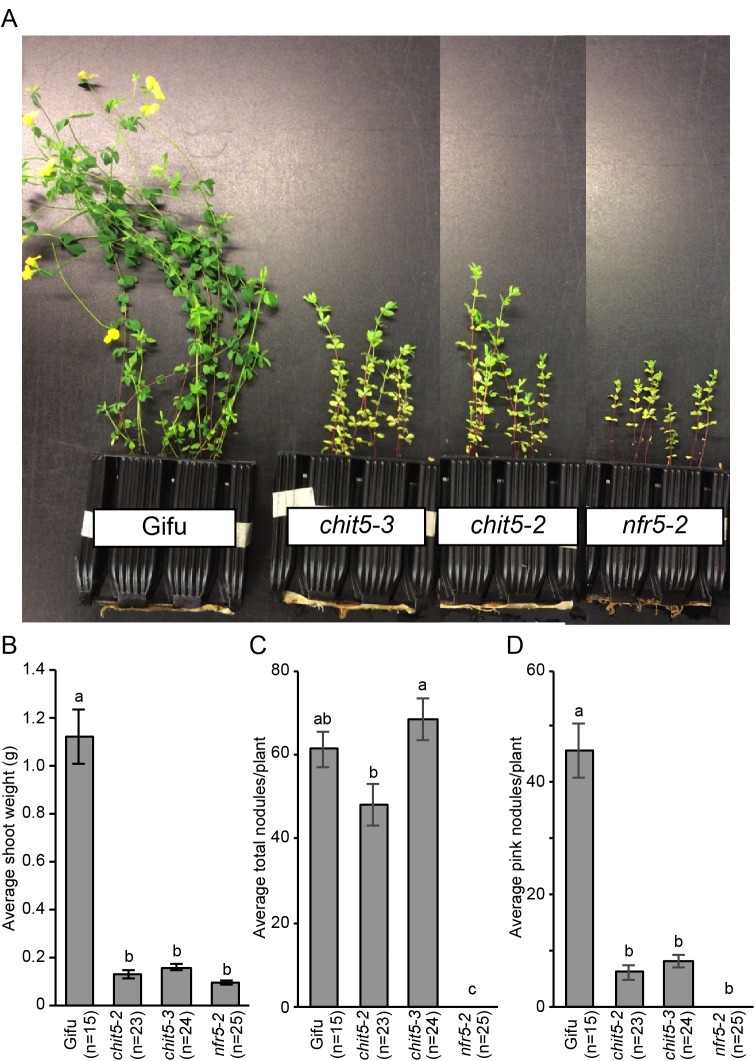
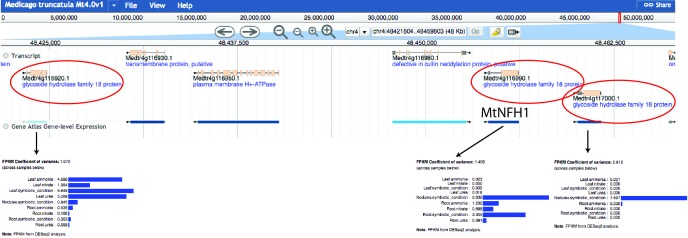
Morphogens provide positional information and their concentration is key to the organized development of multicellular organisms. Nitrogen-fixing root nodules are unique organs induced by Nod factor-producing bacteria. Localized production of Nod factors establishes a developmental field within the root where plant cells are reprogrammed to form infection threads and primordia. We found that regulation of Nod factor levels by is required for the formation of nitrogen-fixing organs, determining the fate of this induced developmental program. Our analysis of plant and bacterial mutants shows that a host chitinase modulates Nod factor levels possibly in a structure-dependent manner. In , this is required for maintaining Nod factor signalling in parallel with the elongation of infection threads within the nodule cortex, while root hair infection and primordia formation are not influenced. Our study shows that infected nodules require balanced levels of Nod factors for completing their transition to functional, nitrogen-fixing organs.
形态发生素提供位置信息,其浓度是多细胞生物有序发育的关键。固氮根瘤是由产生 Nod 因子的细菌诱导的独特器官。Nod 因子的局部产生在根内建立了一个发育场,植物细胞被重新编程形成侵染线和原基。我们发现,通过 调节 Nod 因子水平对于形成固氮器官是必需的,决定了这个诱导发育程序的命运。我们对植物和细菌突变体的分析表明,一种宿主几丁质酶可能以结构依赖的方式调节 Nod 因子水平。在 中,这对于在与侵染线在根瘤皮层内伸长平行的情况下维持 Nod 因子信号是必需的,而根毛侵染和原基形成不受影响。我们的研究表明,感染的根瘤需要平衡的 Nod 因子水平才能完成向功能性固氮器官的转变。