Department of Biological Sciences, St. John's University, New York, NY 11439, USA.
Int J Mol Sci. 2018 Oct 25;19(11):3314. doi: 10.3390/ijms19113314.
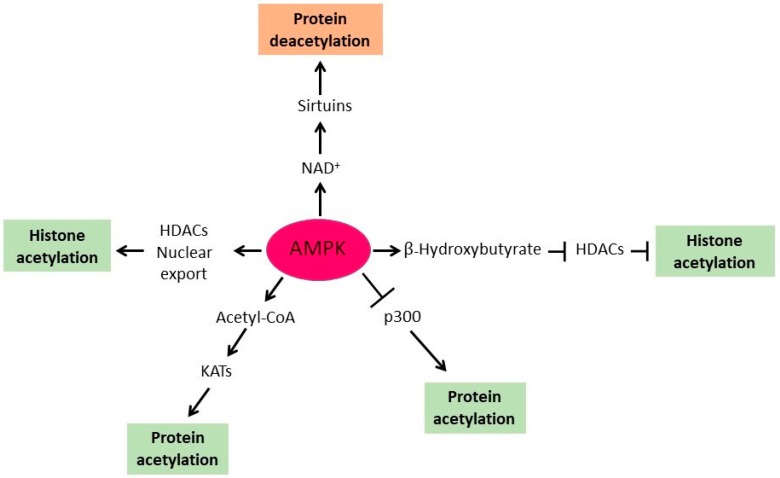
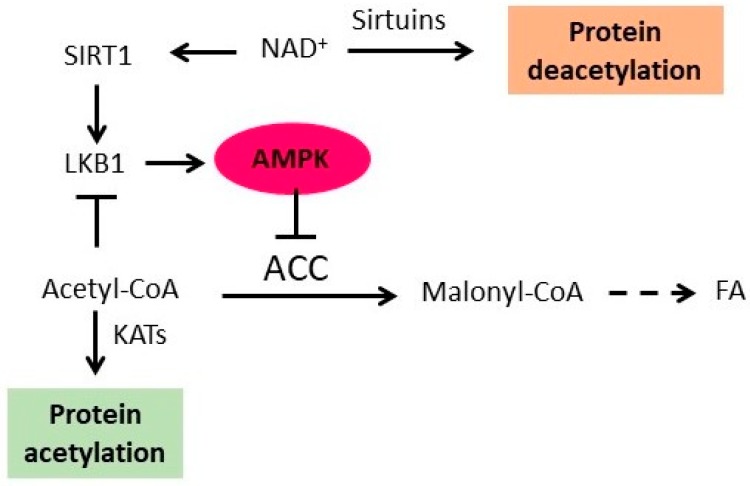
Adenosine monophosphate (AMP)-activated protein kinase (AMPK) serves as an energy sensor and master regulator of metabolism. In general, AMPK inhibits anabolism to minimize energy consumption and activates catabolism to increase ATP production. One of the mechanisms employed by AMPK to regulate metabolism is protein acetylation. AMPK regulates protein acetylation by at least five distinct mechanisms. First, AMPK phosphorylates and inhibits acetyl-CoA carboxylase (ACC) and thus regulates acetyl-CoA homeostasis. Since acetyl-CoA is a substrate for all lysine acetyltransferases (KATs), AMPK affects the activity of KATs by regulating the cellular level of acetyl-CoA. Second, AMPK activates histone deacetylases (HDACs) sirtuins by increasing the cellular concentration of NAD⁺, a cofactor of sirtuins. Third, AMPK inhibits class I and II HDACs by upregulating hepatic synthesis of α-hydroxybutyrate, a natural inhibitor of HDACs. Fourth, AMPK induces translocation of HDACs 4 and 5 from the nucleus to the cytoplasm and thus increases histone acetylation in the nucleus. Fifth, AMPK directly phosphorylates and downregulates p300 KAT. On the other hand, protein acetylation regulates AMPK activity. Sirtuin SIRT1-mediated deacetylation of liver kinase B1 (LKB1), an upstream kinase of AMPK, activates LKB1 and AMPK. AMPK phosphorylates and inactivates ACC, thus increasing acetyl-CoA level and promoting LKB1 acetylation and inhibition. In yeast cells, acetylation of Sip2p, one of the regulatory β-subunits of the SNF1 complex, results in inhibition of SNF1. This results in activation of ACC and reduced cellular level of acetyl-CoA, which promotes deacetylation of Sip2p and activation of SNF1. Thus, in both yeast and mammalian cells, AMPK/SNF1 regulate protein acetylation and are themselves regulated by protein acetylation.
一磷酸腺苷(AMP)激活的蛋白激酶(AMPK)作为能量传感器和代谢的主要调节剂。一般来说,AMPK 抑制合成代谢以最小化能量消耗,并激活分解代谢以增加 ATP 的产生。AMPK 调节代谢的机制之一是蛋白质乙酰化。AMPK 通过至少五种不同的机制调节蛋白质乙酰化。首先,AMPK 磷酸化并抑制乙酰辅酶 A 羧化酶(ACC),从而调节乙酰辅酶 A 的稳态。由于乙酰辅酶 A 是所有赖氨酸乙酰转移酶(KAT)的底物,AMPK 通过调节细胞内乙酰辅酶 A 的水平来影响 KAT 的活性。其次,AMPK 通过增加 NAD⁺的细胞浓度来激活组蛋白去乙酰化酶(HDACs)sirtuins,NAD⁺是 sirtuins 的辅因子。第三,AMPK 通过上调肝合成 α-羟基丁酸来抑制 I 类和 II 类 HDACs,α-羟基丁酸是 HDACs 的天然抑制剂。第四,AMPK 诱导 HDACs 4 和 5 从核内易位到细胞质,从而增加核内组蛋白乙酰化。第五,AMPK 直接磷酸化并下调 p300 KAT。另一方面,蛋白质乙酰化调节 AMPK 活性。SIRT1 介导的 AMPK 的上游激酶肝激酶 B1(LKB1)的去乙酰化激活 LKB1 和 AMPK。AMPK 磷酸化并失活 ACC,从而增加乙酰辅酶 A 水平,促进 LKB1 乙酰化和抑制。在酵母细胞中,SNF1 复合物的调节β亚基之一 Sip2p 的乙酰化导致 SNF1 的抑制。这导致 ACC 的激活和细胞内乙酰辅酶 A 水平的降低,促进 Sip2p 的去乙酰化和 SNF1 的激活。因此,在酵母和哺乳动物细胞中,AMPK/SNF1 调节蛋白质乙酰化,并且自身受蛋白质乙酰化调节。