Institute of Molecular Biology, University of Oregon, Eugene, United States.
Department of Biochemistry Faculty of Science, Charles University, Prague, Czech Republic.
Elife. 2018 Nov 23;7:e41497. doi: 10.7554/eLife.41497.
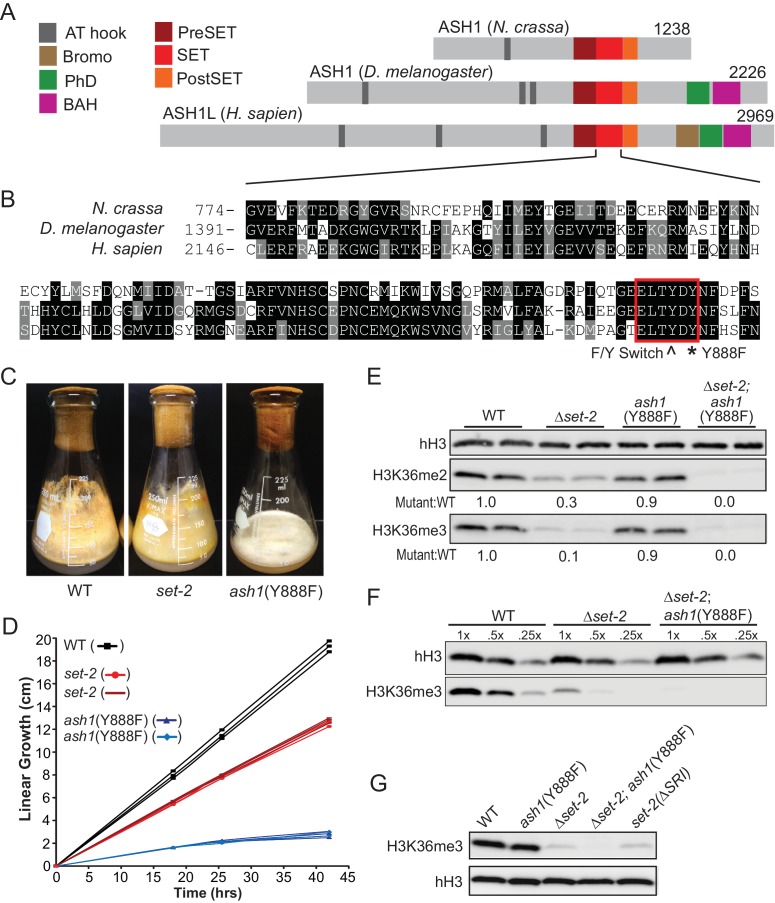
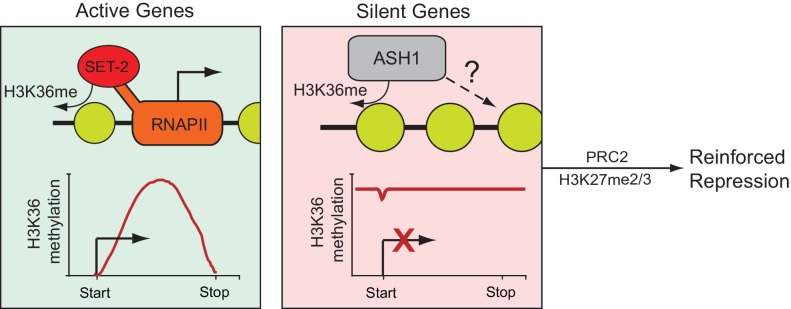
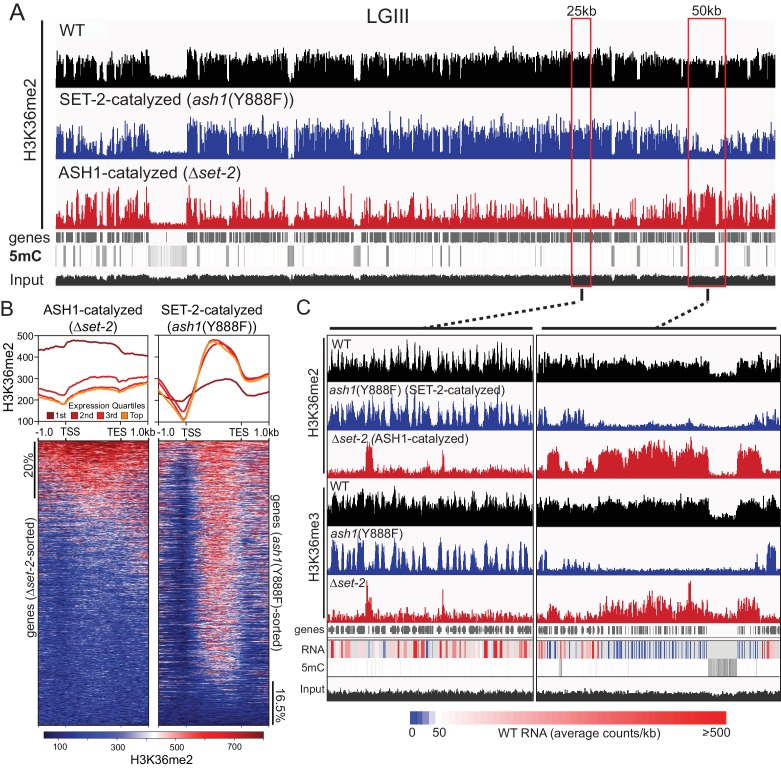
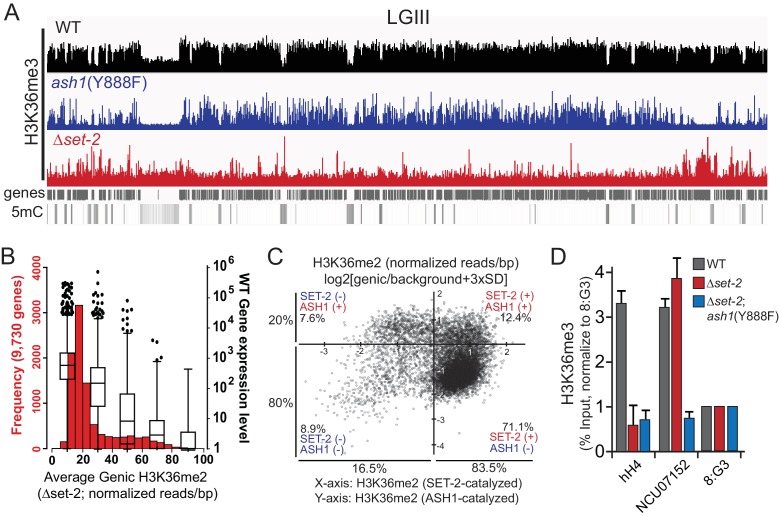
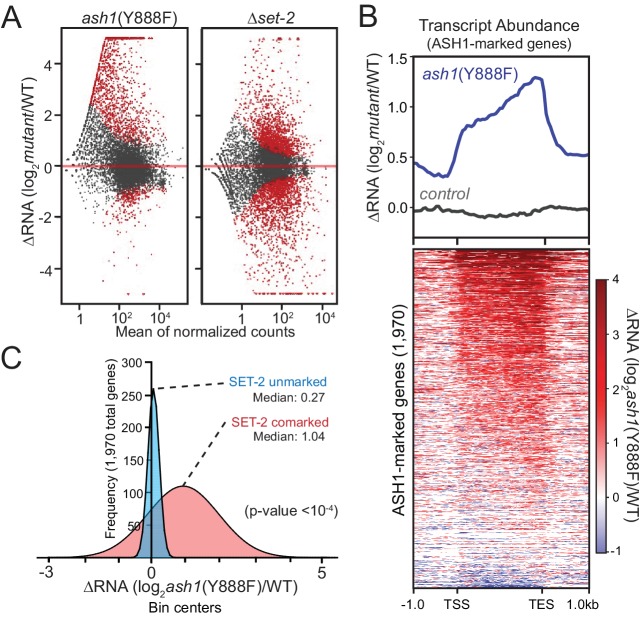
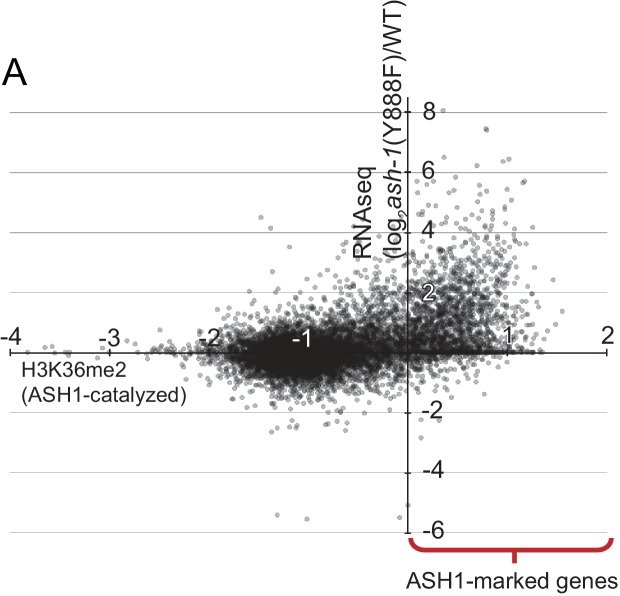
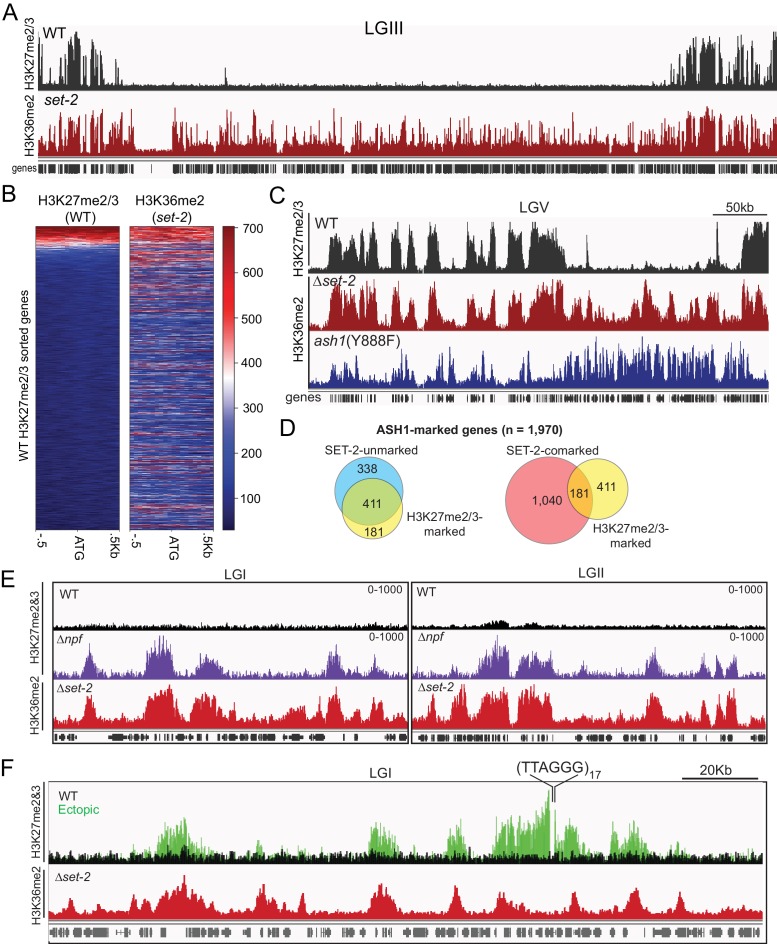
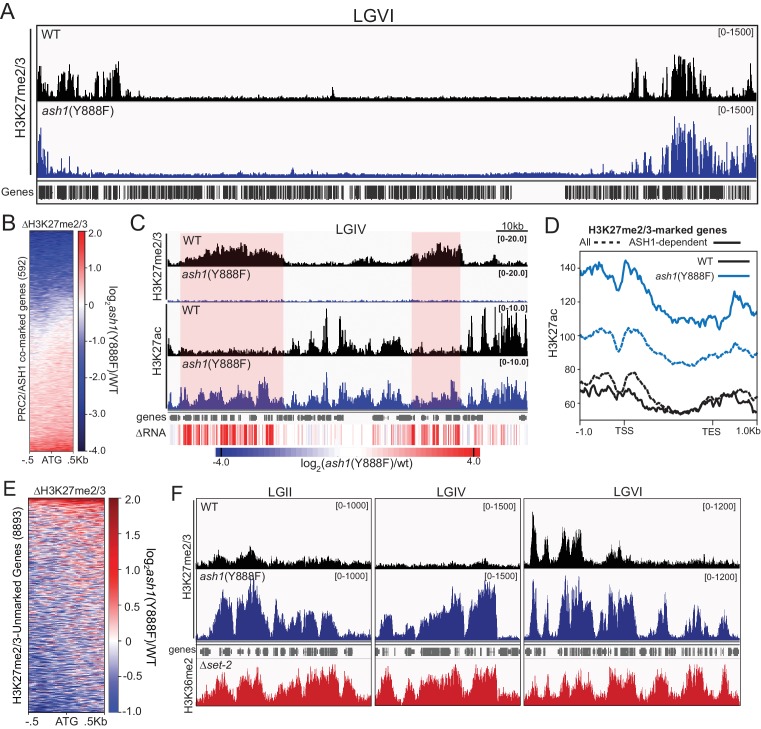
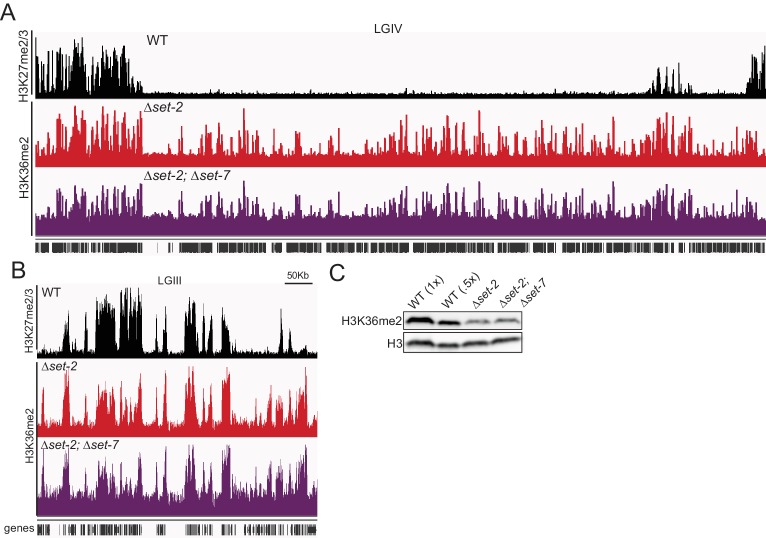
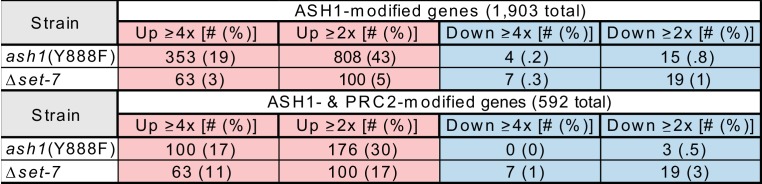
Methylation of histone H3 at lysine 36 (H3K36me), a widely-distributed chromatin mark, largely results from association of the lysine methyltransferase (KMT) SET-2 with RNA polymerase II (RNAPII), but most eukaryotes also have additional H3K36me KMTs that act independently of RNAPII. These include the orthologs of ASH1, which are conserved in animals, plants, and fungi but whose function and control are poorly understood. We found that has just two H3K36 KMTs, ASH1 and SET-2, and were able to explore the function and distribution of each enzyme independently. While H3K36me deposited by SET-2 marks active genes, inactive genes are modified by ASH1 and its activity is critical for their repression. ASH1-marked chromatin can be further modified by methylation of H3K27, and ASH1 catalytic activity modulates the accumulation of H3K27me2/3 both positively and negatively. These findings provide new insight into ASH1 function, H3K27me2/3 establishment, and repression in facultative heterochromatin.
组蛋白 H3 赖氨酸 36 位的甲基化(H3K36me)是一种广泛分布的染色质标记,主要源于赖氨酸甲基转移酶(KMT)SET-2 与 RNA 聚合酶 II(RNAPII)的结合,但大多数真核生物也有其他独立于 RNAPII 的 H3K36me KMT。这些包括在动物、植物和真菌中保守的 ASH1 的同源物,但它们的功能和调控机制了解甚少。我们发现,酵母只含有两种 H3K36 KMT,ASH1 和 SET-2,并且能够独立探索每种酶的功能和分布。虽然 SET-2 沉积的 H3K36me 标记活性基因,但非活性基因由 ASH1 修饰,其活性对于它们的抑制至关重要。ASH1 标记的染色质可以进一步被 H3K27 的甲基化修饰,并且 ASH1 的催化活性对 H3K27me2/3 的积累既有正向作用也有负向作用。这些发现为 ASH1 功能、H3K27me2/3 的建立以及兼性异染色质的抑制提供了新的见解。