Institute of Molecular Biology, University of Oregon, Eugene, United States.
Faculty of Medical Sciences, University of Fukui, Fukui, Japan.
Elife. 2018 Jan 3;7:e31216. doi: 10.7554/eLife.31216.
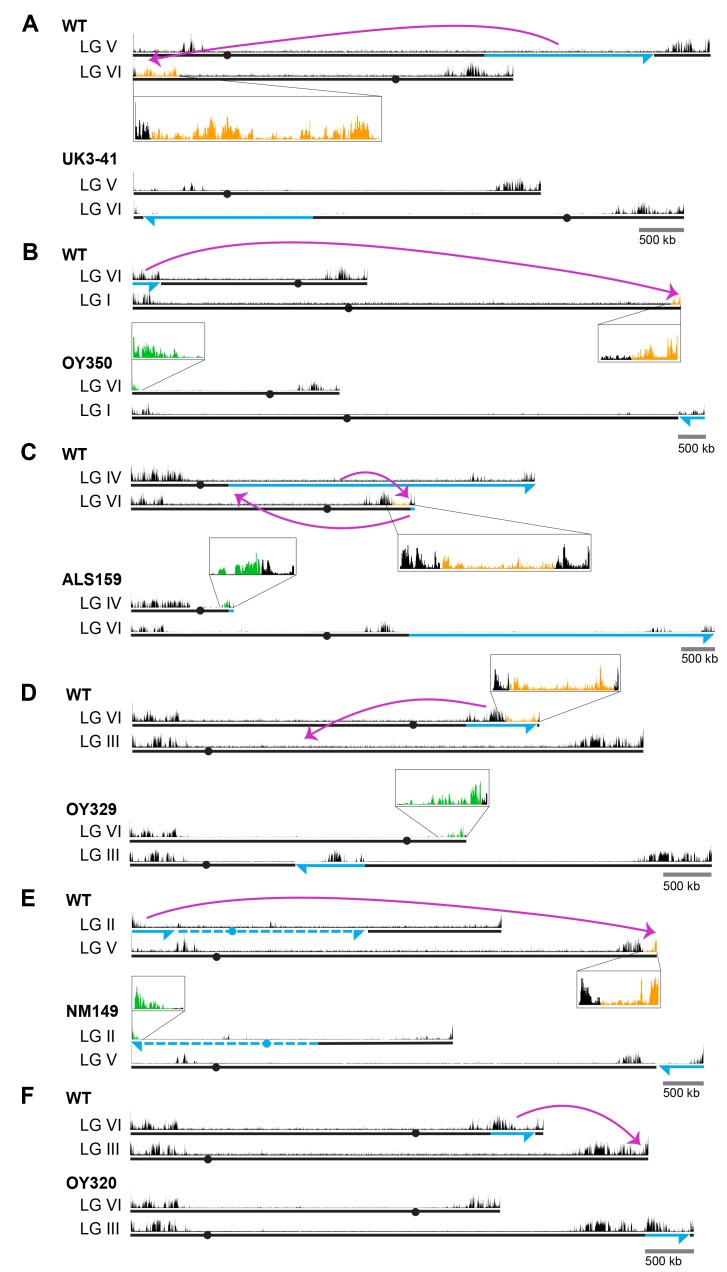
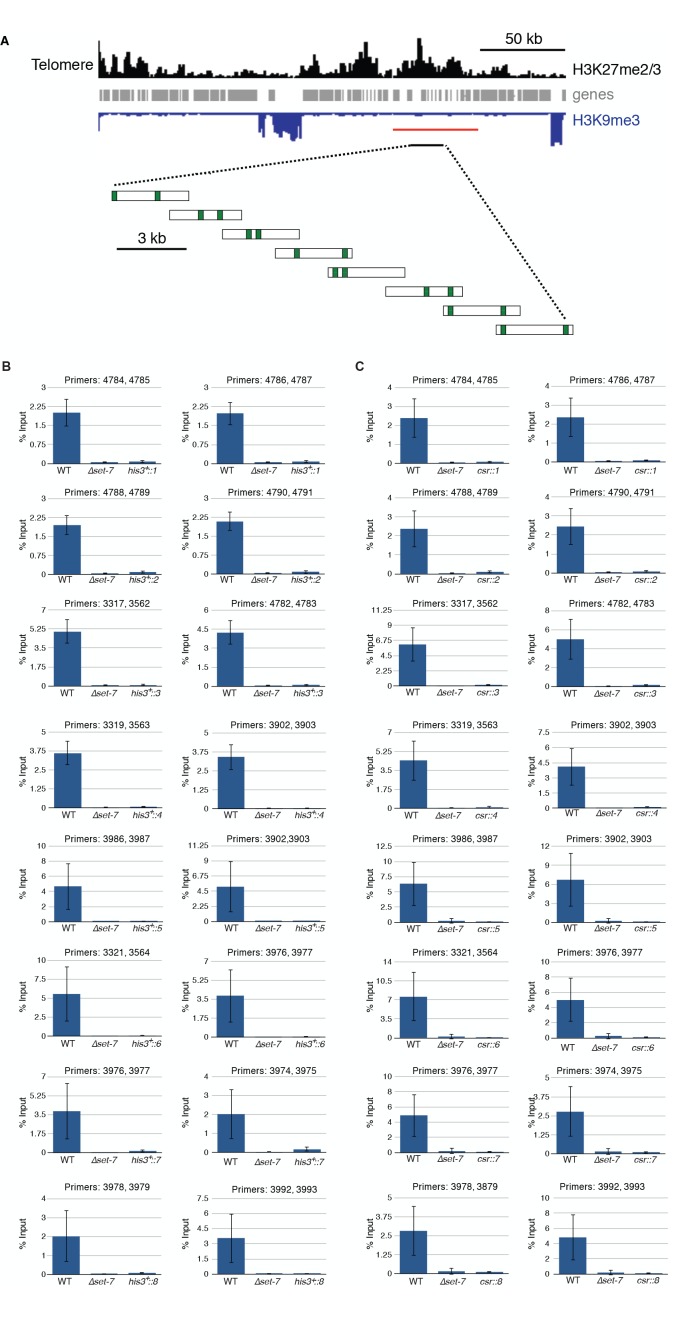
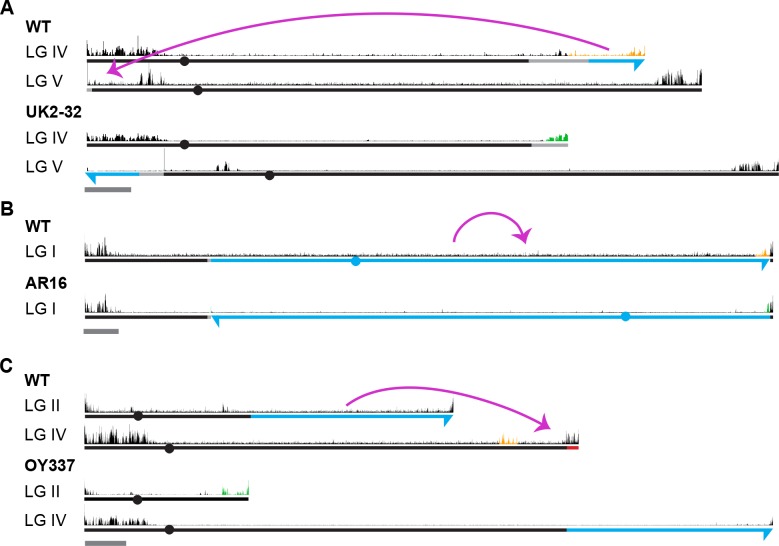
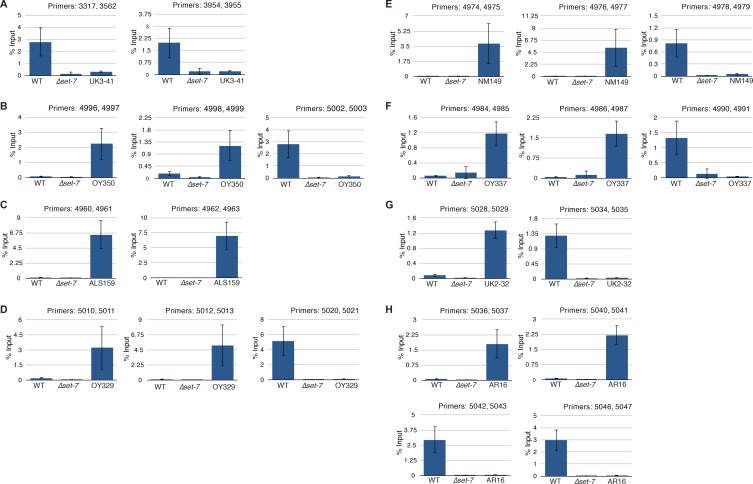
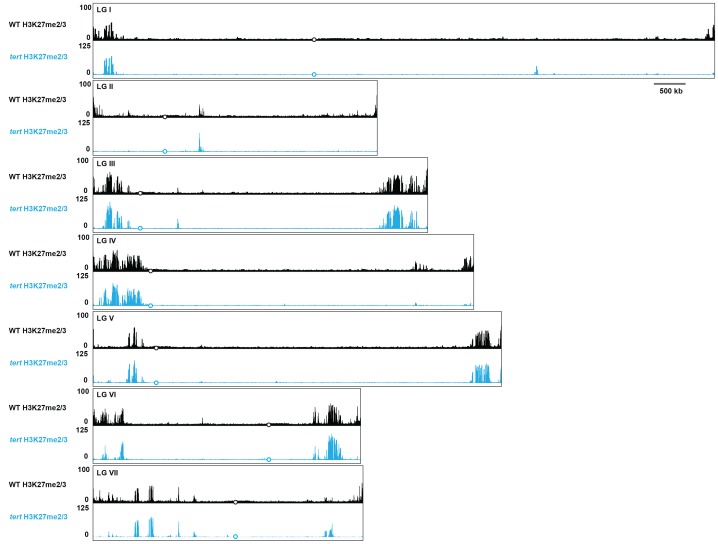
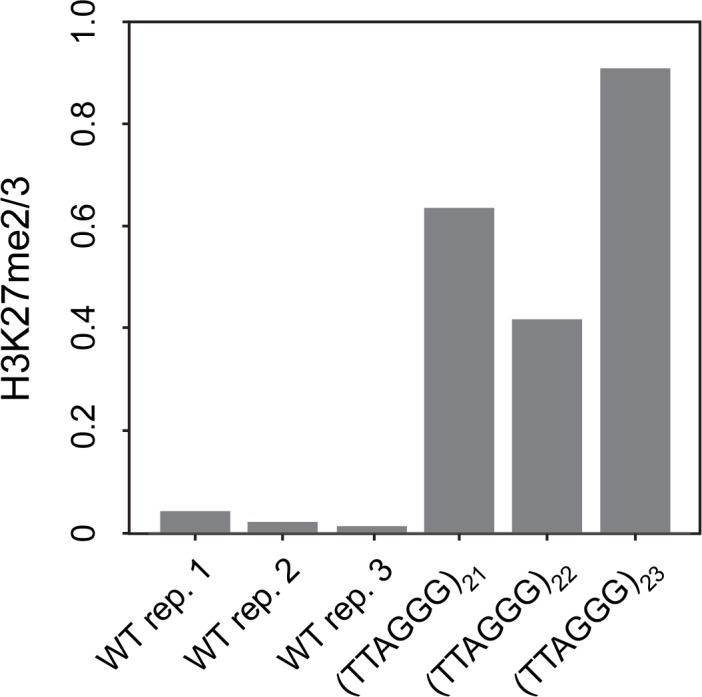
Development in higher organisms requires selective gene silencing, directed in part by di-/trimethylation of lysine 27 on histone H3 (H3K27me2/3). Knowledge of the cues that control formation of such repressive Polycomb domains is extremely limited. We exploited natural and engineered chromosomal rearrangements in the fungus to elucidate the control of H3K27me2/3. Analyses of H3K27me2/3 in strains bearing chromosomal rearrangements revealed both position-dependent and position-independent facultative heterochromatin. We found that proximity to chromosome ends is necessary to maintain, and sufficient to induce, transcriptionally repressive, subtelomeric H3K27me2/3. We ascertained that such telomere-proximal facultative heterochromatin requires native telomere repeats and found that a short array of ectopic telomere repeats, (TTAGGG) can induce a large domain (~225 kb) of H3K27me2/3. This provides an example of a -acting sequence that directs H3K27 methylation. Our findings provide new insight into the relationship between genome organization and control of heterochromatin formation.
高等生物的发育需要有选择性的基因沉默,这部分是由组蛋白 H3 赖氨酸 27 的二甲基化/三甲基化(H3K27me2/3)来指导的。然而,控制这种抑制性多梳域形成的线索知识极为有限。我们利用真菌中的天然和工程染色体重排来阐明 H3K27me2/3 的控制。对携带染色体重排的菌株中的 H3K27me2/3 进行分析,揭示了位置依赖性和位置非依赖性的兼性异染色质。我们发现,接近染色体末端对于维持和诱导转录抑制性端粒异染色质是必要的。我们确定了这种端粒近端的兼性异染色质需要天然端粒重复序列,并发现短的异位端粒重复序列(TTAGGG)可以诱导 H3K27me2/3 的大片段(~225 kb)。这提供了一个 - 作用序列指导 H3K27 甲基化的例子。我们的发现为基因组组织和异染色质形成控制之间的关系提供了新的见解。