Protein Processing Section, Structural Biophysics Laboratory, Center for Cancer Research, National Cancer Institute, Frederick, MD 21702, USA.
Protein Processing Section, Structural Biophysics Laboratory, Center for Cancer Research, National Cancer Institute, Frederick, MD 21702, USA; Linganore High School, Frederick, MD 21701, USA.
J Mol Biol. 2019 Mar 1;431(5):939-955. doi: 10.1016/j.jmb.2019.01.021. Epub 2019 Jan 18.
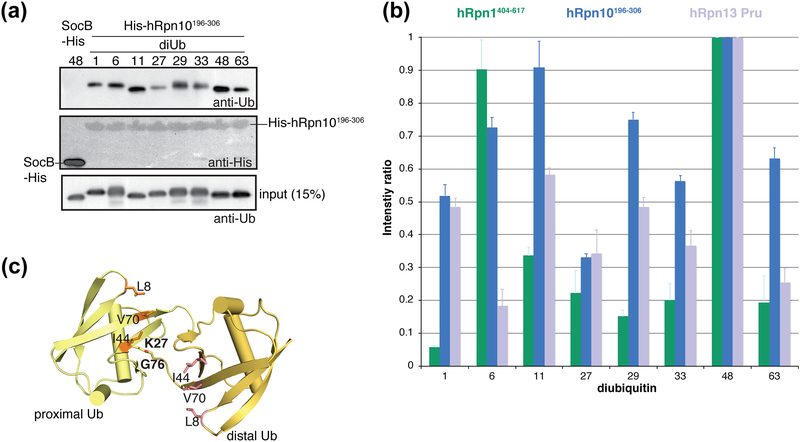
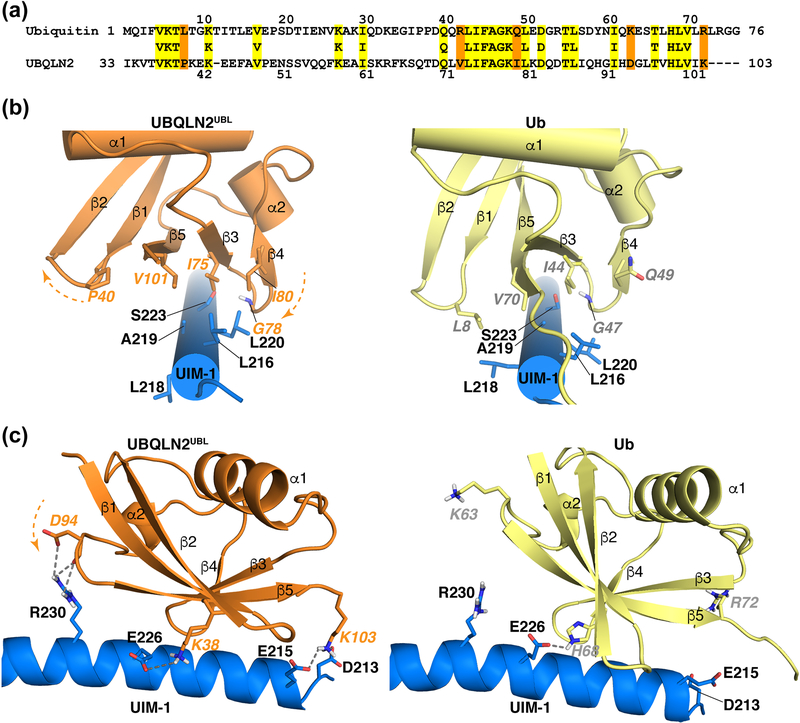
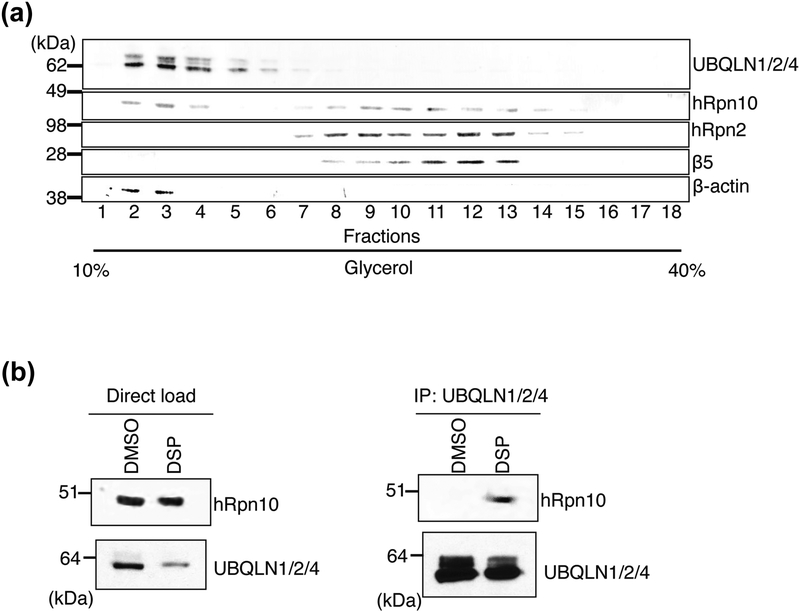
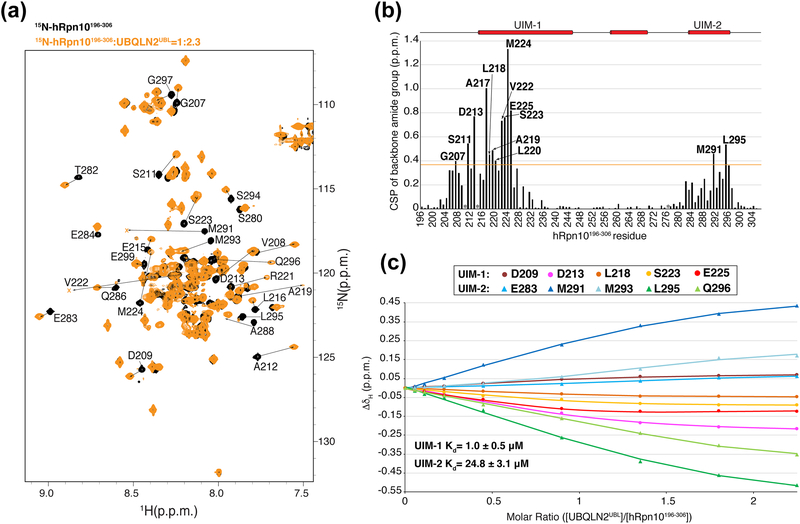
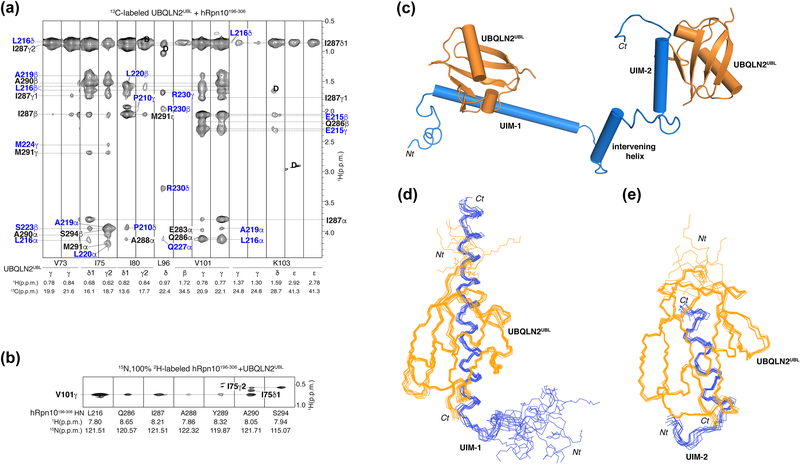
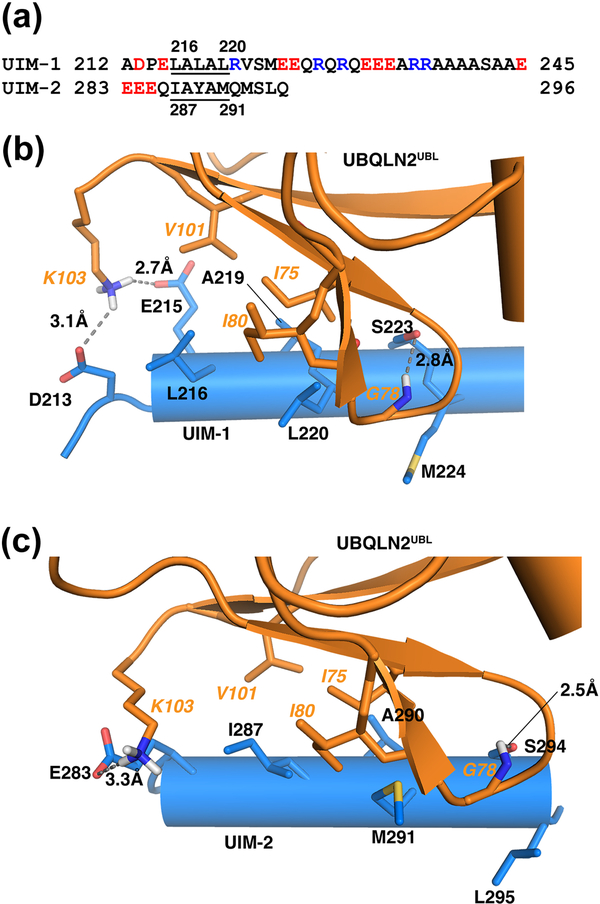
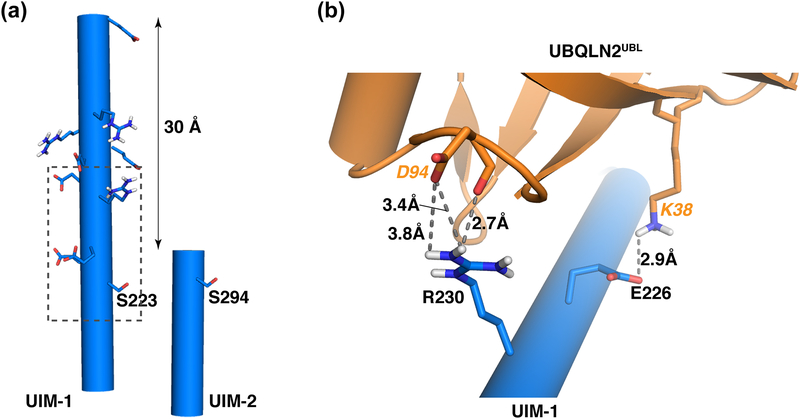
The 26S proteasome is a highly complex 2.5-MDa molecular machine responsible for regulated protein degradation. Proteasome substrates are typically marked by ubiquitination for recognition at receptor sites contributed by Rpn1/S2/PSMD2, Rpn10/S5a, and Rpn13/Adrm1. Each receptor site can bind substrates directly by engaging conjugated ubiquitin chains or indirectly by binding to shuttle factors Rad23/HR23, Dsk2/PLIC/UBQLN, or Ddi1, which contain a ubiquitin-like domain (UBL) that adopts the ubiquitin fold. Previous structural studies have defined how each of the proteasome receptor sites binds to ubiquitin chains as well as some of the interactions that occur with the shuttle factors. Here, we define how hRpn10 binds to the UBQLN2 UBL domain, solving the structure of this complex by NMR, and determine affinities for each UIM region by a titration experiment. UBQLN2 UBL exhibits 25-fold stronger affinity for the N-terminal UIM-1 over UIM-2 of hRpn10. Moreover, we discover that UBQLN2 UBL is fine-tuned for the hRpn10 UIM-1 site over the UIM-2 site by taking advantage of the additional contacts made available through the longer UIM-1 helix. We also test hRpn10 versatility for the various ubiquitin chains to find less specificity for any particular linkage type compared to hRpn1 and hRpn13, as expected from the flexible linker region that connects the two UIMs; nonetheless, hRpn10 does exhibit some preference for K48 and K11 linkages. Altogether, these results provide new insights into the highly complex and complementary roles of the proteasome receptor sites and shuttle factors.
26S 蛋白酶体是一种高度复杂的 2.5MDa 分子机器,负责调节蛋白质降解。蛋白酶体底物通常通过泛素化标记,以便在由 Rpn1/S2/PSMD2、Rpn10/S5a 和 Rpn13/Adrm1 贡献的受体部位被识别。每个受体部位可以通过直接结合缀合的泛素链或间接结合穿梭因子 Rad23/HR23、Dsk2/PLIC/UBQLN 或 Ddi1 来识别底物,这些穿梭因子包含一个泛素样结构域 (UBL),该结构域采用泛素折叠。以前的结构研究已经定义了蛋白酶体受体部位如何结合泛素链以及与穿梭因子发生的一些相互作用。在这里,我们通过 NMR 确定了 hRpn10 与 UBQLN2 UBL 结构域的结合方式,并通过滴定实验确定了每个 UIM 区域的亲和力。UBQLN2 UBL 对 hRpn10 的 N 端 UIM-1 的亲和力比对 UIM-2 的亲和力强 25 倍。此外,我们发现 UBQLN2 UBL 通过利用通过较长的 UIM-1 螺旋提供的额外接触,针对 hRpn10 的 UIM-1 位点进行了微调,而不是 UIM-2 位点。我们还测试了 hRpn10 对各种泛素链的多功能性,与预期的一样,由于连接两个 UIM 的柔性连接区,与 hRpn1 和 hRpn13 相比,它对任何特定的连接类型的特异性较低;尽管如此,hRpn10 确实对 K48 和 K11 连接表现出一些偏好。总的来说,这些结果为蛋白酶体受体部位和穿梭因子的高度复杂和互补作用提供了新的见解。