Biovision team, Université Côte d'Azur, Inria, France.
INPHYNI, Université Côte d'Azur, CNRS, Sophia Antipolis, France.
Sci Rep. 2019 Feb 12;9(1):1859. doi: 10.1038/s41598-018-38299-4.
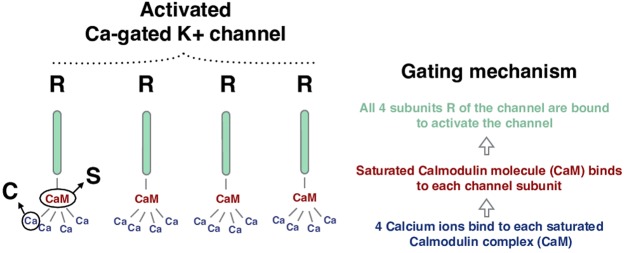
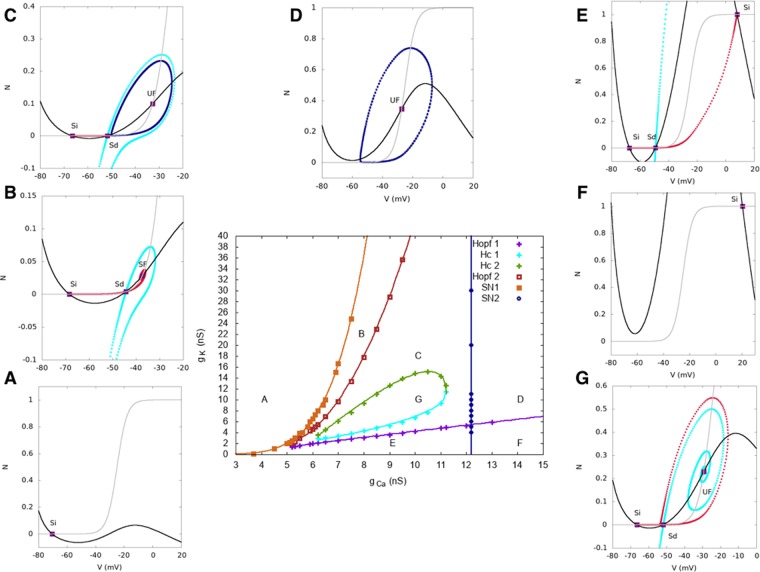
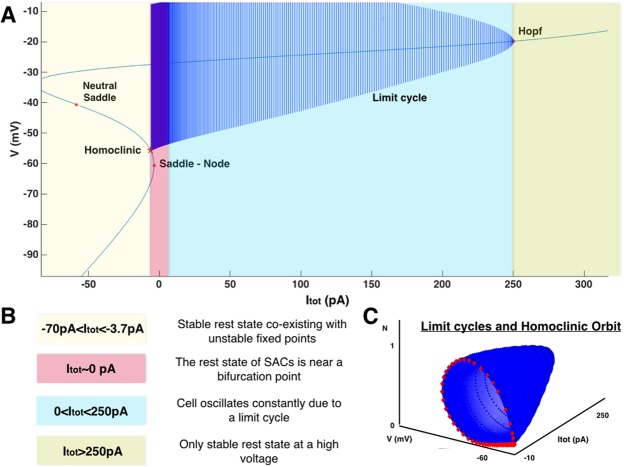
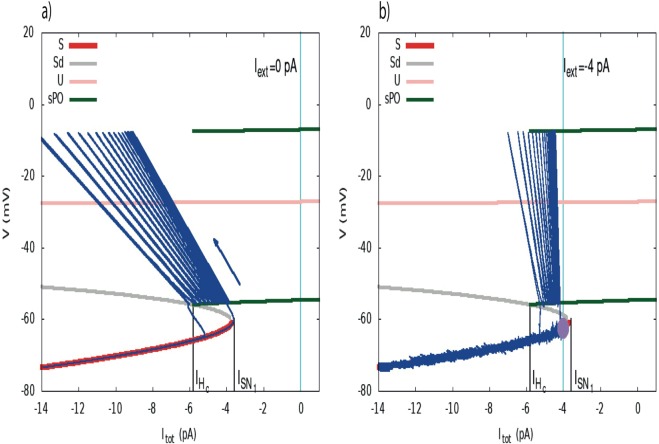
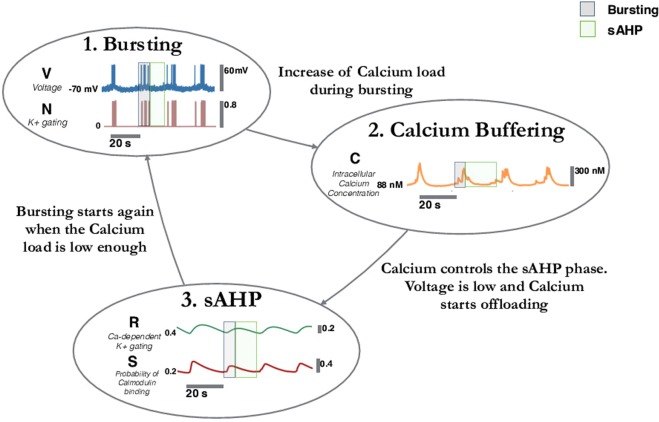
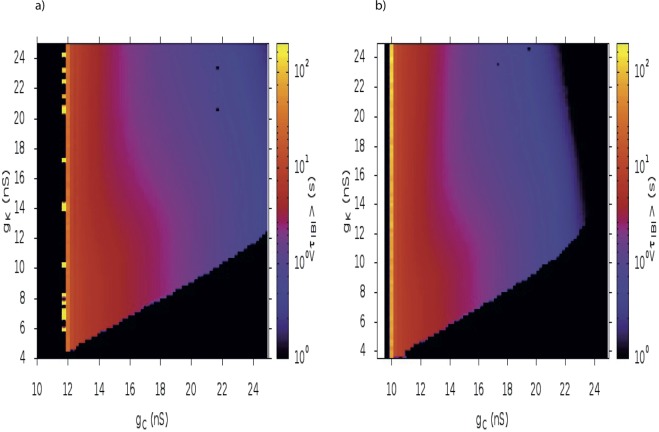
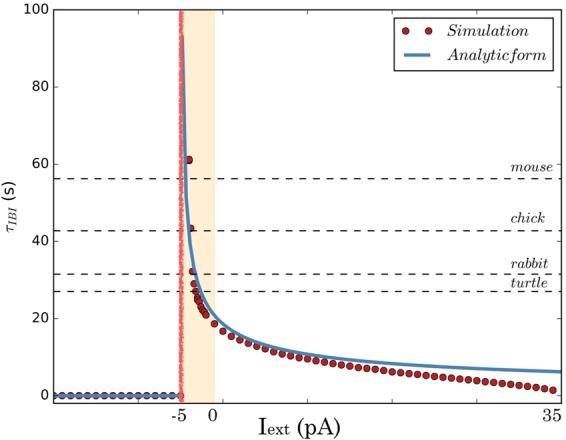
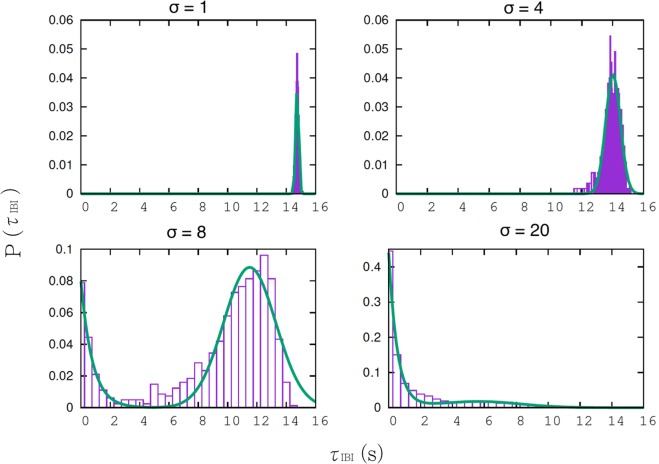
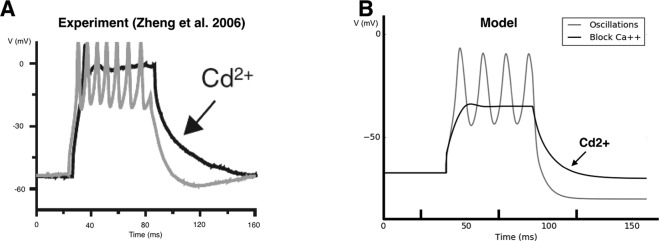
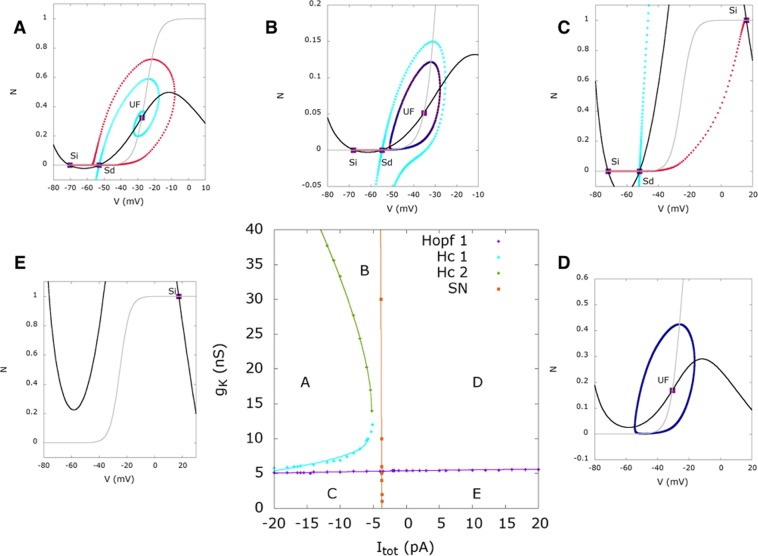
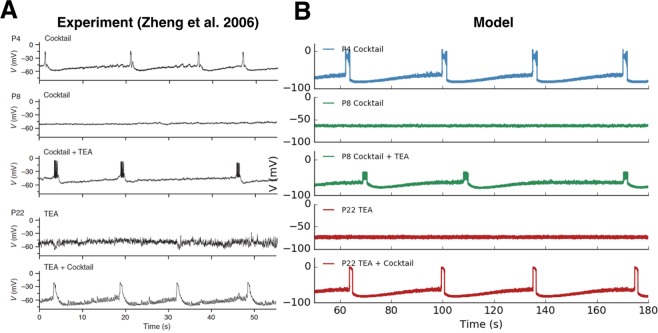
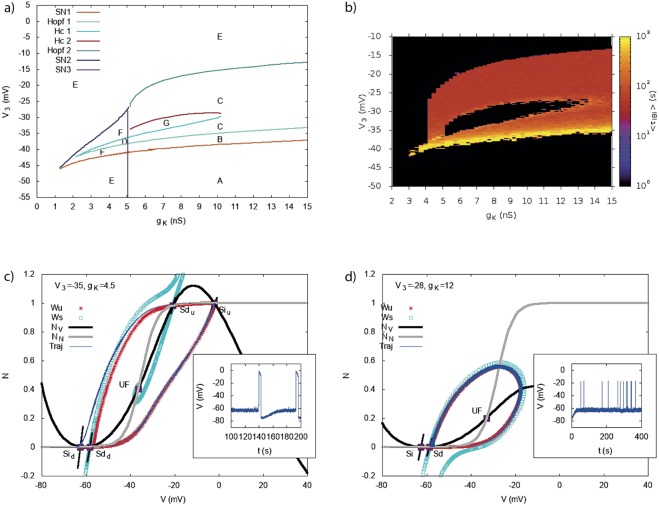
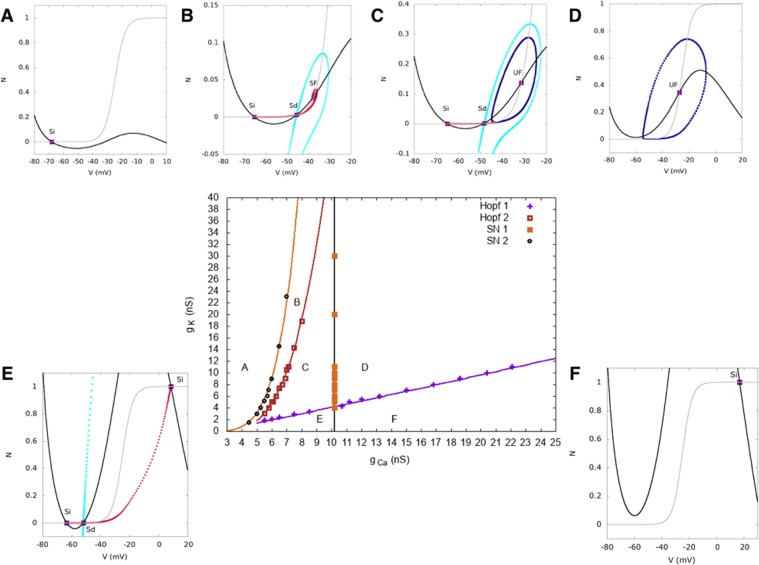
During early development, waves of activity propagate across the retina and play a key role in the proper wiring of the early visual system. During a particular phase of the retina development (stage II) these waves are triggered by a transient network of neurons, called Starburst Amacrine Cells (SACs), showing a bursting activity which disappears upon further maturation. The underlying mechanisms of the spontaneous bursting and the transient excitability of immature SACs are not completely clear yet. While several models have attempted to reproduce retinal waves, none of them is able to mimic the rhythmic autonomous bursting of individual SACs and reveal how these cells change their intrinsic properties during development. Here, we introduce a mathematical model, grounded on biophysics, which enables us to reproduce the bursting activity of SACs and to propose a plausible, generic and robust, mechanism that generates it. The core parameters controlling repetitive firing are fast depolarizing V-gated calcium channels and hyperpolarizing V-gated potassium channels. The quiescent phase of bursting is controlled by a slow after hyperpolarization (sAHP), mediated by calcium-dependent potassium channels. Based on a bifurcation analysis we show how biophysical parameters, regulating calcium and potassium activity, control the spontaneously occurring fast oscillatory activity followed by long refractory periods in individual SACs. We make a testable experimental prediction on the role of voltage-dependent potassium channels on the excitability properties of SACs and on the evolution of this excitability along development. We also propose an explanation on how SACs can exhibit a large variability in their bursting periods, as observed experimentally within a SACs network as well as across different species, yet based on a simple, unique, mechanism. As we discuss, these observations at the cellular level have a deep impact on the retinal waves description.
在早期发育过程中,活动波在视网膜中传播,并在早期视觉系统的正确布线中发挥关键作用。在视网膜发育的特定阶段(阶段 II),这些波是由称为星爆型无长突细胞(Starburst Amacrine Cells,SAC)的短暂神经元网络触发的,这些细胞表现出爆发活动,随着进一步成熟而消失。自发性爆发和不成熟 SACs 的短暂兴奋性的潜在机制尚不完全清楚。虽然已经有几个模型试图复制视网膜波,但没有一个能够模拟单个 SAC 的节律性自主爆发,并揭示这些细胞在发育过程中如何改变其内在特性。在这里,我们引入了一个基于生物物理学的数学模型,该模型使我们能够复制 SAC 的爆发活动,并提出一个合理、通用且稳健的机制来产生这种活动。控制重复放电的核心参数是快速去极化 V 门控钙通道和超极化 V 门控钾通道。爆发的静息期由钙依赖性钾通道介导的慢后超极化(slow after hyperpolarization,sAHP)控制。基于分岔分析,我们展示了调节钙和钾活性的生物物理参数如何控制单个 SACs 中自发发生的快速振荡活动,随后是长时间的不应期。我们对电压依赖性钾通道在 SACs 兴奋性特性上的作用以及这种兴奋性在发育过程中的演变提出了可测试的实验预测。我们还提出了一种解释,说明 SACs 如何表现出其爆发周期的大变化,正如在 SACs 网络以及不同物种中观察到的那样,但基于一种简单、独特的机制。正如我们所讨论的,这些细胞水平的观察对视网膜波的描述有深远的影响。