Institute for Biophysics, University of Bremen, Otto-Hahn-Allee 1, 28359 Bremen, Germany.
PLoS One. 2019 Apr 23;14(4):e0215622. doi: 10.1371/journal.pone.0215622. eCollection 2019.
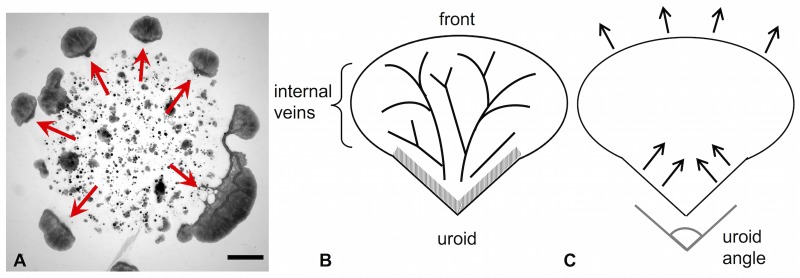
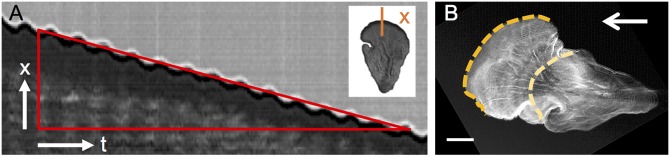
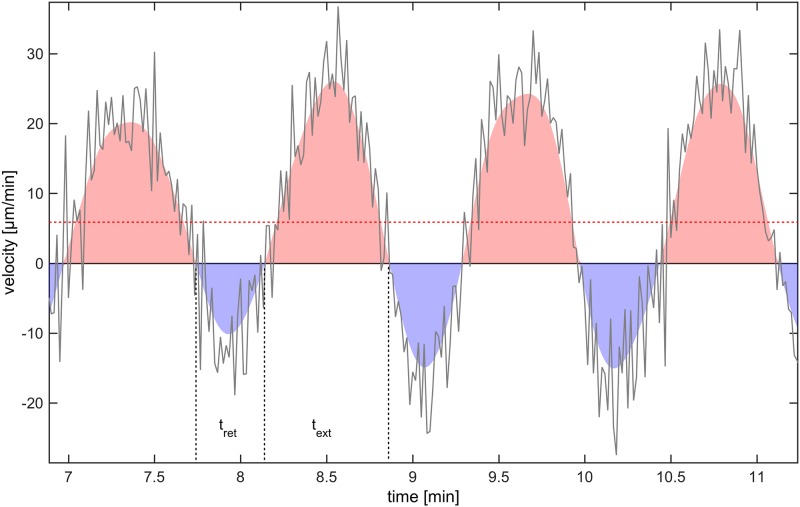
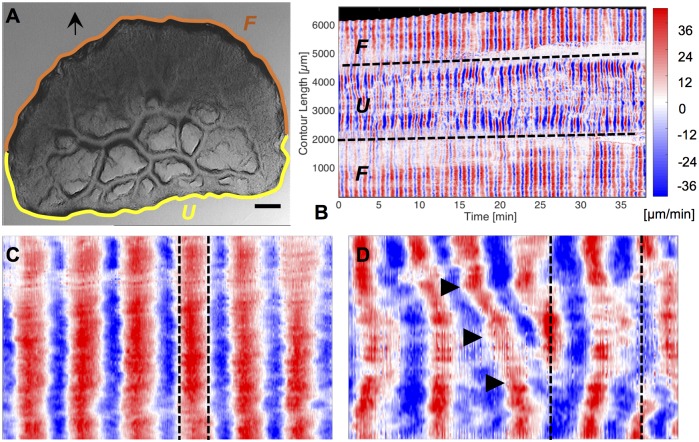
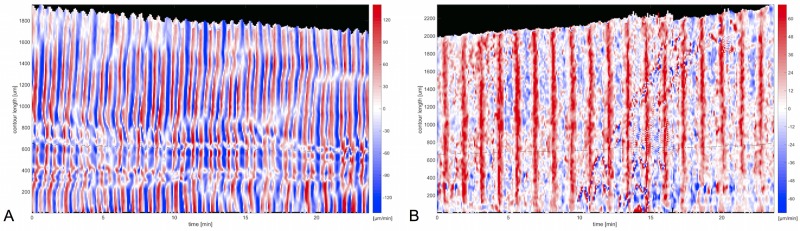
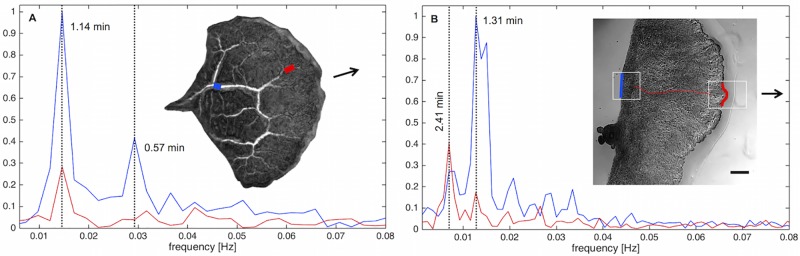
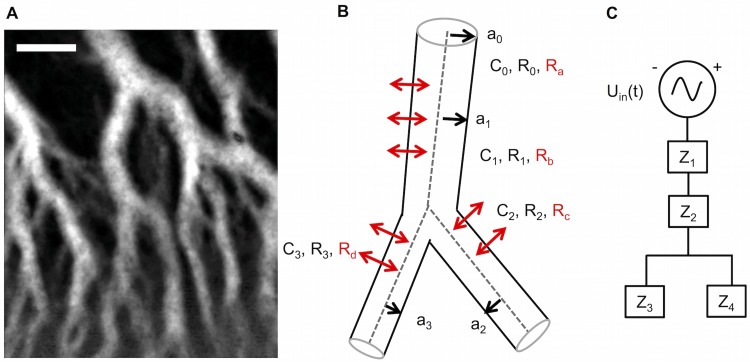
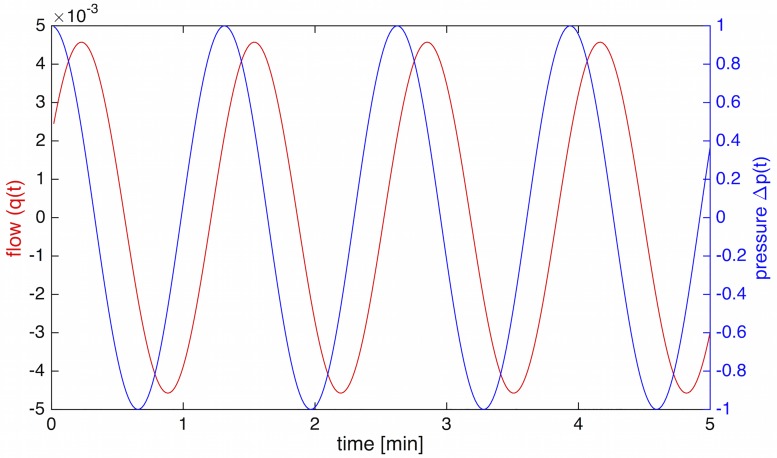
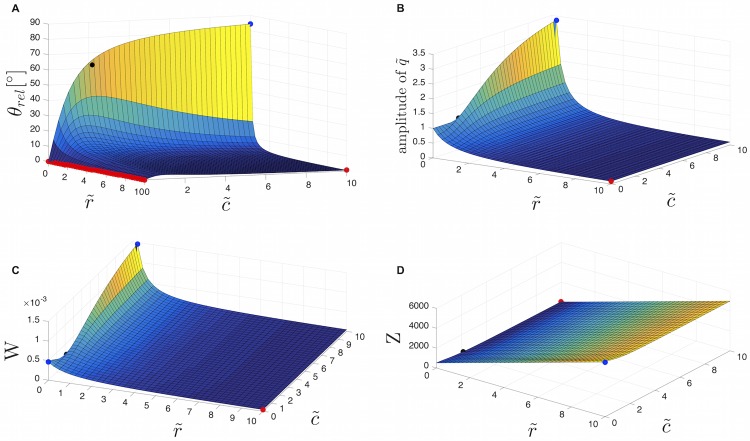
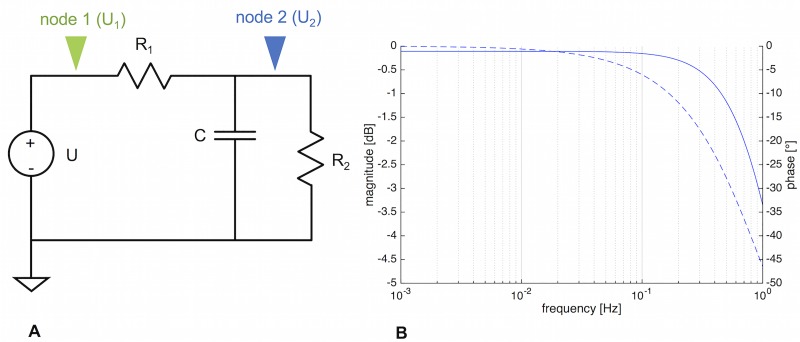
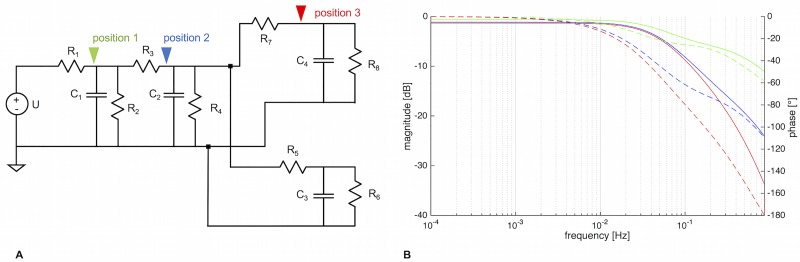
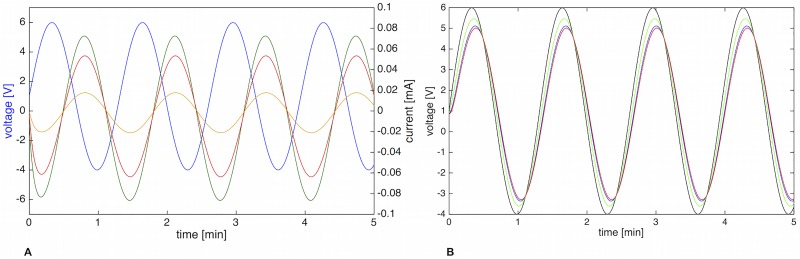
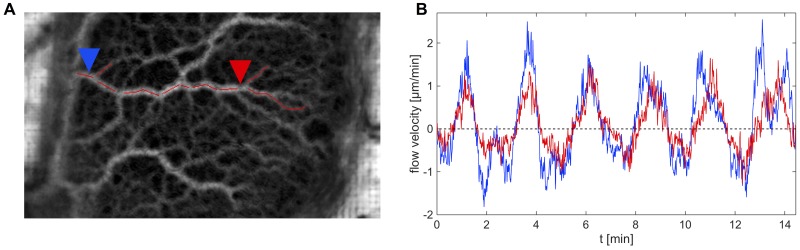
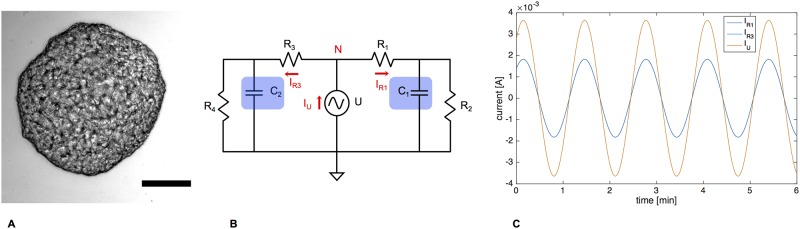
The plasmodial slime mold Physarum polycephalum exhibits strong, periodic flow of cytoplasm through the veins of its network. In the special case of mesoplasmodia, a newly described starvation-induced, shape-constant morphotype, this periodic endoplasm streaming is the basis of locomotion. Furthermore, we presume that cytoplasm flow is also involved in signal transmission and signal processing. Mesoplasmodia motility resembles amoeboid locomotion. In contrast to other amoebae, however, mesoplasmodia move without extending pseudopods and retain a coherent, fan-shaped morphology throughout their steady locomotion. Attaining sizes of up to 2 mm2, mesoplasmodia are also much bigger than other amoebae. We characterize this particular type of locomotion and identify patterns of movement. By using the analogy between pulsatile fluid flow through a network of elastic tubes and electrical circuits, we build a lumped model that explains observed fluid flow patterns. Essentially, the mesoplasmodium acts as a low-pass filter, permitting only low-frequency oscillations to propagate from back to front. This frequency selection serves to optimize flow and reduces power dissipation. Furthermore, we introduce a distributed element into the lumped model to explain cell polarization during the onset of chemotaxis: Biochemical cues (internal or external) lead to a local softening of the actin cortex, which in turn causes an increased flow of cytoplasm into that area and, thus, a net forward movement. We conclude that the internal actin-enclosed vein network gives the slime mold a high measure of control over fluid transport, especially by softening or hardening, which in turn leads to polarization and net movement.
多头绒泡菌的原生质团表现出强烈的、周期性的细胞质流动,通过其网络的脉管。在新描述的饥饿诱导的、形状恒定的形态型mesoplasmodia 的特殊情况下,这种周期性的内质体流动是运动的基础。此外,我们推测细胞质流动也参与信号传输和信号处理。mesoplasmodia 的运动类似于变形虫运动。然而,与其他变形虫不同的是,mesoplasmodia 在没有延伸伪足的情况下移动,并在其稳定运动过程中保持连贯的扇形形态。mesoplasmodia 可以达到 2 平方毫米的大小,也比其他变形虫大得多。我们描述了这种特殊类型的运动并确定了运动模式。通过将脉冲式流体流过弹性管网络与电路进行类比,我们构建了一个集中模型,解释了观察到的流体流动模式。本质上,mesoplasmodia 充当低通滤波器,只允许低频振荡从前向后传播。这种频率选择有助于优化流动并减少功率损耗。此外,我们在集中模型中引入了一个分布式元件,以解释趋化作用开始时的细胞极化:生化线索(内部或外部)导致肌动蛋白皮层局部软化,这反过来又导致细胞质向该区域的流动增加,从而导致净向前运动。我们得出的结论是,内部的肌动蛋白封闭的脉管网赋予粘菌对流体运输的高度控制,特别是通过软化或硬化,这反过来又导致极化和净运动。