Department of Structural Biology, University of Pittsburgh School of Medicine, Pittsburgh, PA 15260.
Department of Pharmacology and Chemical Biology, University of Pittsburgh School of Medicine, Pittsburgh, PA 15260.
Proc Natl Acad Sci U S A. 2019 May 7;116(19):9333-9339. doi: 10.1073/pnas.1814999116. Epub 2019 Apr 24.
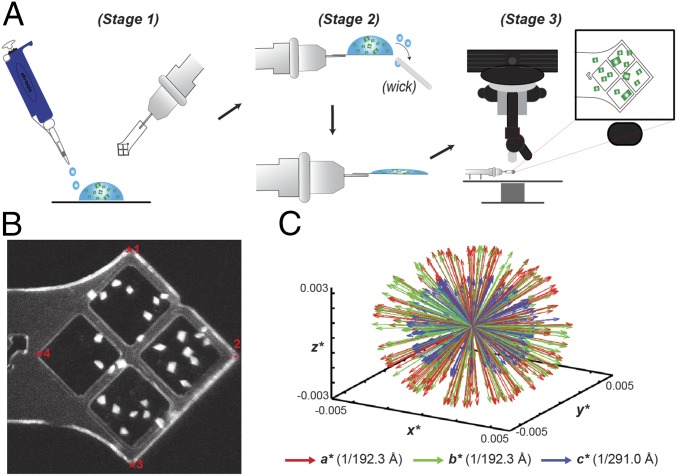
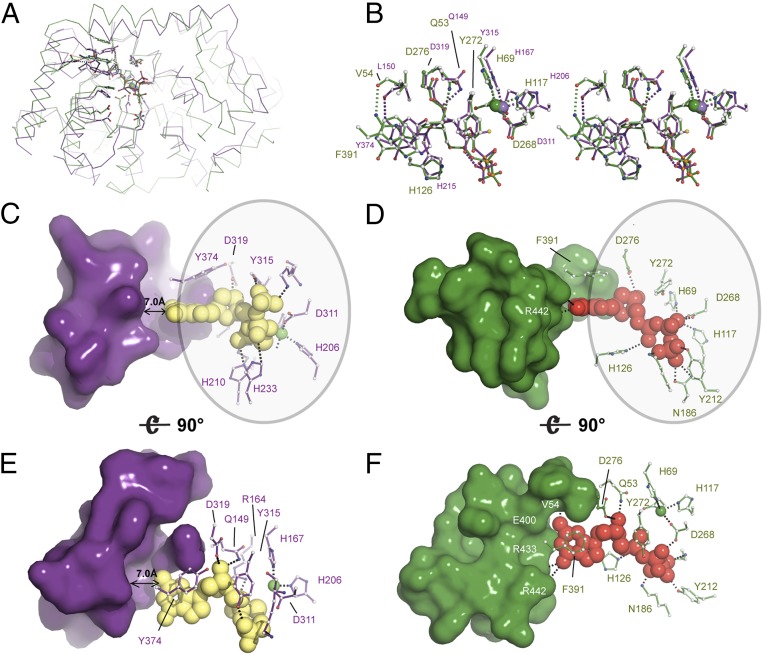
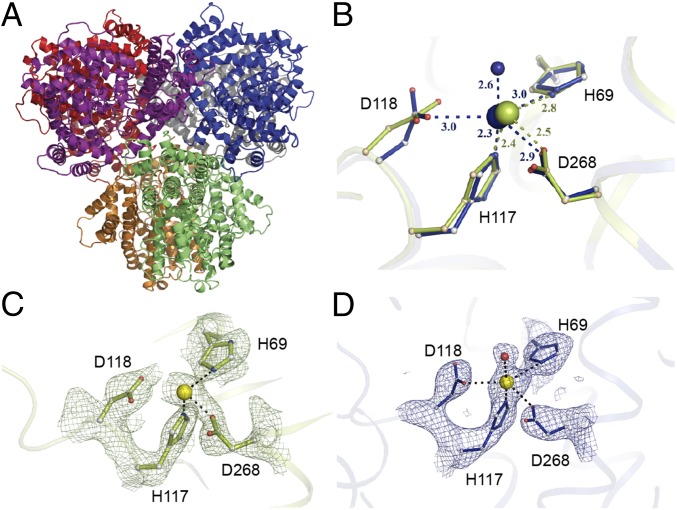
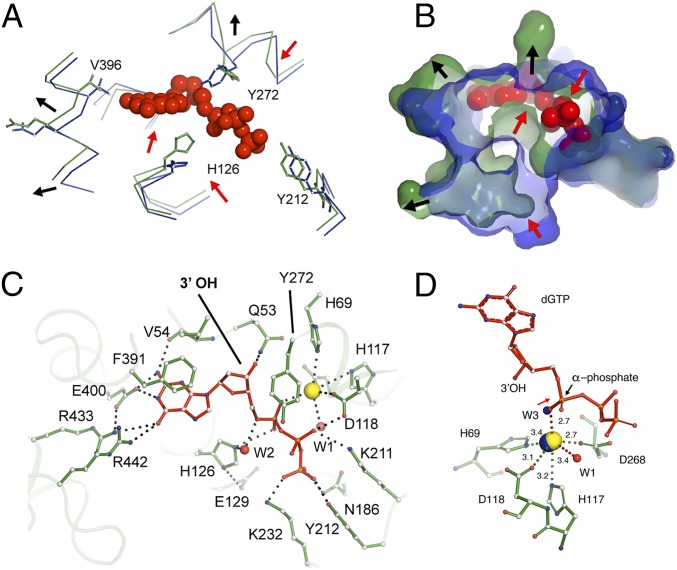
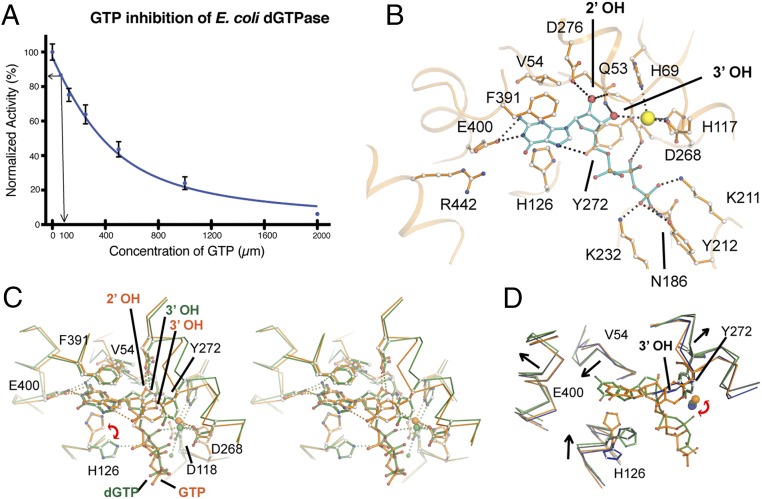
Deoxynucleotide triphosphohydrolases (dNTPases) play a critical role in cellular survival and DNA replication through the proper maintenance of cellular dNTP pools. While the vast majority of these enzymes display broad activity toward canonical dNTPs, such as the dNTPase SAMHD1 that blocks reverse transcription of retroviruses in macrophages by maintaining dNTP pools at low levels, ()dGTPase is the only known enzyme that specifically hydrolyzes dGTP. However, the mechanism behind dGTP selectivity is unclear. Here we present the free-, ligand (dGTP)- and inhibitor (GTP)-bound structures of hexameric dGTPase, including an X-ray free-electron laser structure of the free -dGTPase enzyme to 3.2 Å. To obtain this structure, we developed a method that applied UV-fluorescence microscopy, video analysis, and highly automated goniometer-based instrumentation to map and rapidly position individual crystals randomly located on fixed target holders, resulting in the highest indexing rates observed for a serial femtosecond crystallography experiment. Our structures show a highly dynamic active site where conformational changes are coupled to substrate (dGTP), but not inhibitor binding, since GTP locks dGTPase in its apo- form. Moreover, despite no sequence homology, -dGTPase and SAMHD1 share similar active-site and HD motif architectures; however, -dGTPase residues at the end of the substrate-binding pocket mimic Watson-Crick interactions providing guanine base specificity, while a 7-Å cleft separates SAMHD1 residues from dNTP bases, abolishing nucleotide-type discrimination. Furthermore, the structures shed light on the mechanism by which long distance binding (25 Å) of single-stranded DNA in an allosteric site primes the active site by conformationally "opening" a tyrosine gate allowing enhanced substrate binding.
脱氧核苷酸三磷酸水解酶(dNTPases)通过适当维持细胞内的 dNTP 池,在细胞存活和 DNA 复制中发挥着至关重要的作用。虽然这些酶中的绝大多数对典型的 dNTP 具有广泛的活性,例如 SAMHD1 这种酶通过将 dNTP 池维持在低水平来阻断巨噬细胞中的逆转录病毒的逆转录,()但 dGTPase 是唯一已知的专门水解 dGTP 的酶。然而,dGTP 选择性的机制尚不清楚。在这里,我们展示了六聚体 dGTPase 的游离态、配体(dGTP)结合态和抑制剂(GTP)结合态结构,包括自由态 dGTPase 的 X 射线自由电子激光结构,分辨率达到 3.2Å。为了获得这个结构,我们开发了一种方法,该方法应用紫外荧光显微镜、视频分析和高度自动化的基于测角仪的仪器,对随机位于固定靶标上的单个晶体进行定位和快速定位,从而获得了串联飞秒结晶学实验中观察到的最高索引率。我们的结构显示了一个高度动态的活性位点,其中构象变化与底物(dGTP)偶联,但与抑制剂结合不偶联,因为 GTP 将 dGTPase 锁定在其无配体形式。此外,尽管没有序列同源性,但-β-dGTPase 和 SAMHD1 具有相似的活性位点和 HD 基序结构;然而,β-dGTPase 位于底物结合口袋末端的残基模拟 Watson-Crick 相互作用,提供鸟嘌呤碱基特异性,而 SAMHD1 残基与 dNTP 碱基之间有 7Å 的裂隙,消除了核苷酸类型的辨别。此外,这些结构揭示了远距离结合(25Å)的单链 DNA 在别构位点通过构象“打开”酪氨酸门,从而增强底物结合的机制,从而使活性位点被“开启”。