Laboratory of Molecular Pathology, Department of Health Sciences, Università del Piemonte Orientale "A. Avogadro", Novara, Italy.
Present address: Zuckerberg San Francisco General Hospital and Trauma Center, University of California San Francisco, San Francisco, CA, 94110, USA.
Cell Commun Signal. 2019 May 2;17(1):39. doi: 10.1186/s12964-019-0354-2.
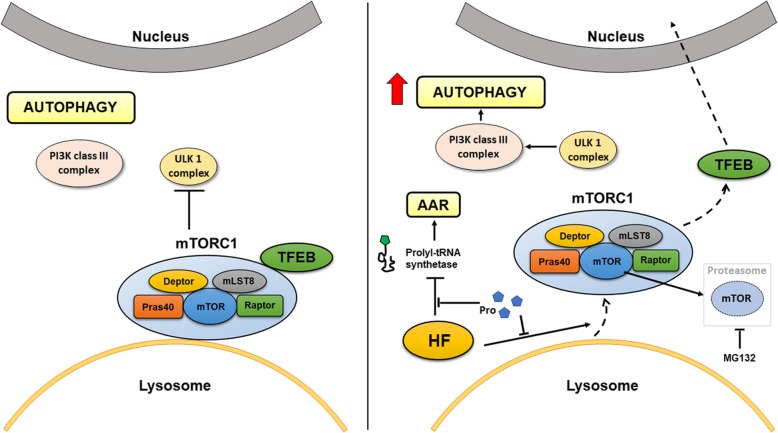
In the event of amino acid starvation, the cell activates two main protective pathways: Amino Acid starvation Response (AAR), to inhibit global translation, and autophagy, to recover the essential substrates from degradation of redundant self-components. Whether and how AAR and autophagy (ATG) are cross-regulated and at which point the two regulatory pathways intersect remain unknown. Here, we provide experimental evidence that the mammalian target of rapamycin (mTOR) complex 1 (mTORC1) specifically located at the lysosome level links the AAR with the autophagy pathway.
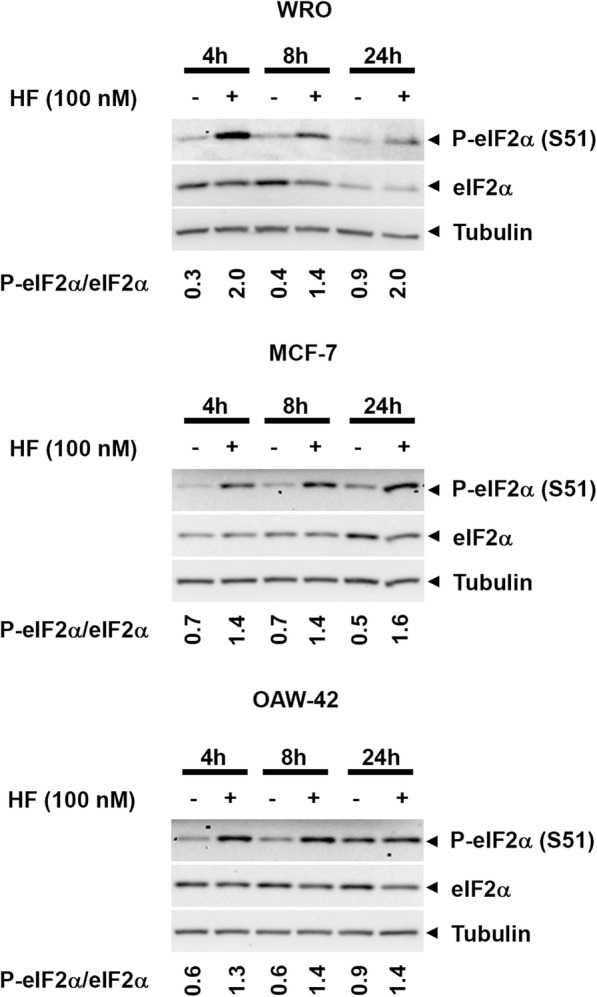
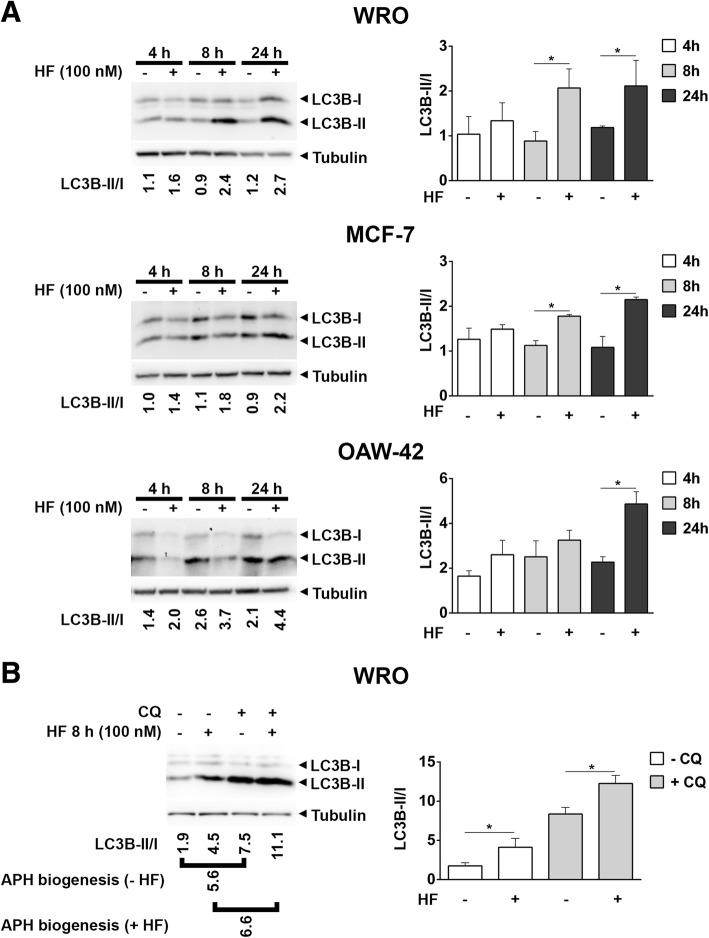
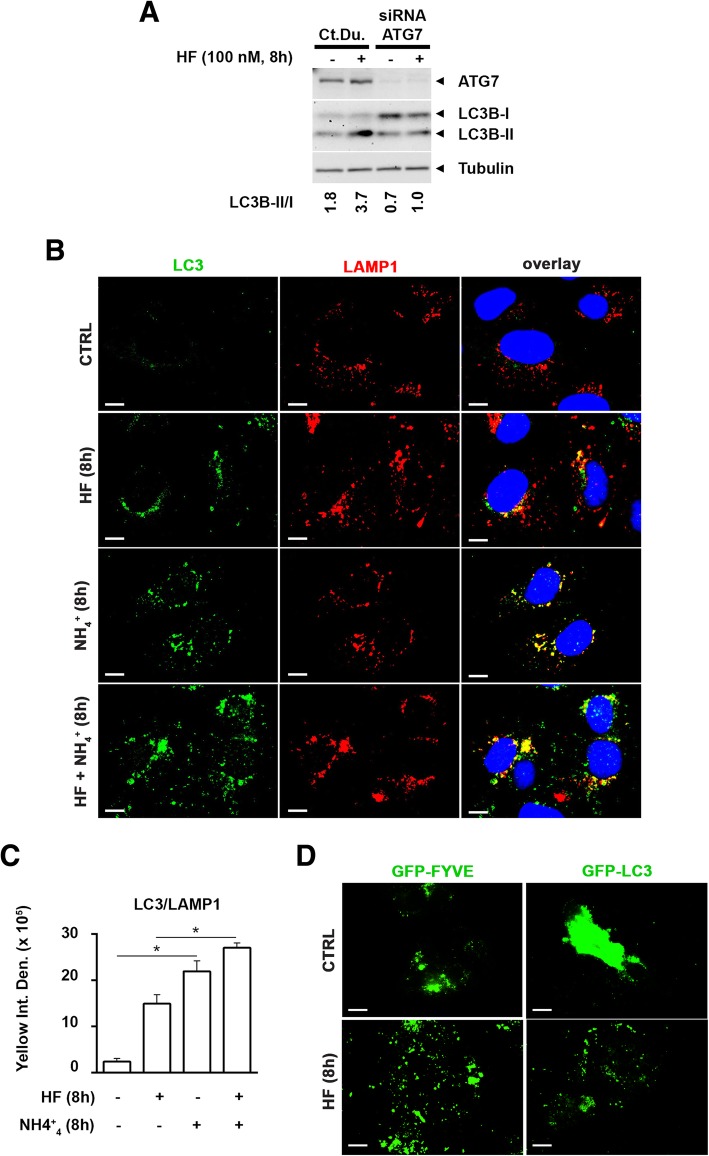
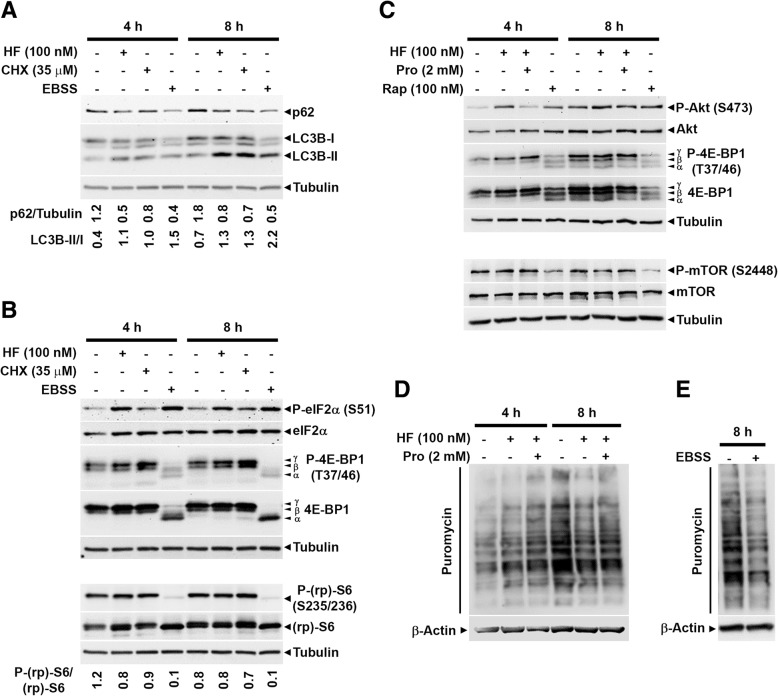
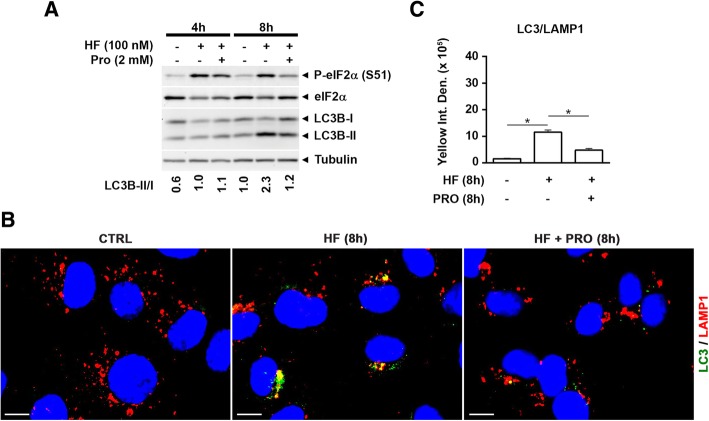
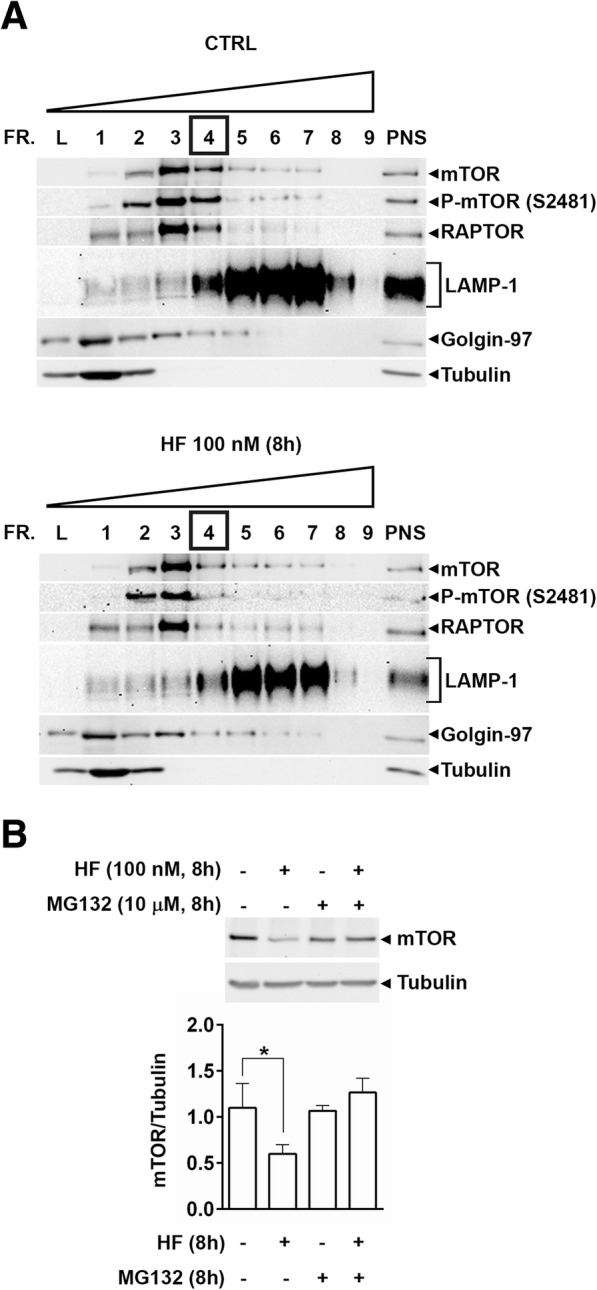
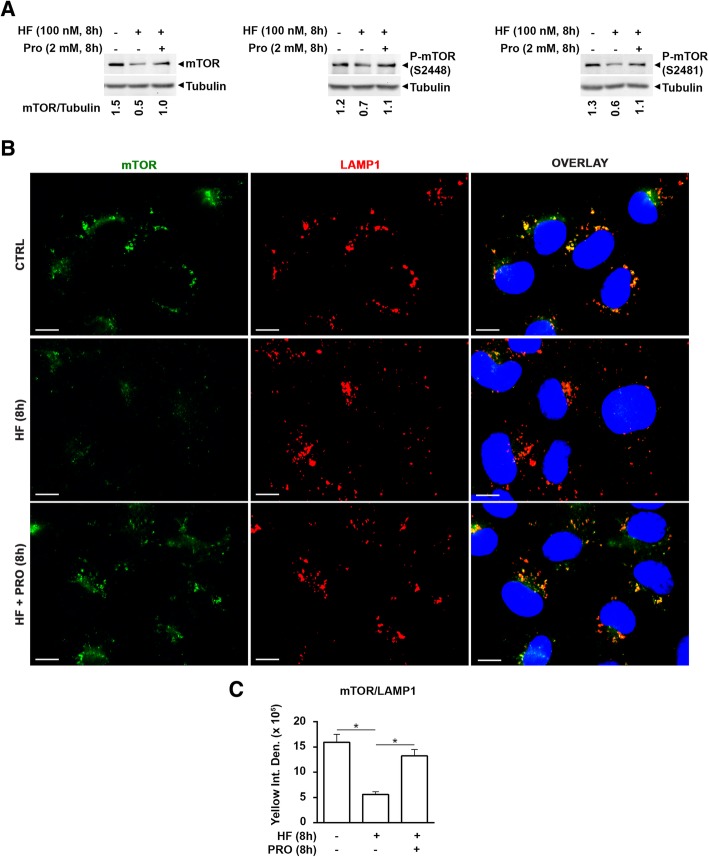
As an inducer of the AAR, we used halofuginone (HF), an alkaloid that binds to the prolyl-tRNA synthetase thus mimicking the unavailability of proline (PRO). Induction of AAR was determined assessing the phosphorylation of the eukaryotic translation initiation factor (eIF) 2α. Autophagy was monitored by assessing the processing and accumulation of microtubule-associated protein 1 light chain 3 isoform B (LC3B) and sequestosome-1 (p62/SQSTM1) levels. The activity of mTORC1 was monitored through assessment of the phosphorylation of mTOR, (rp)S6 and 4E-BP1. Global protein synthesis was determined by puromycin incorporation assay. mTORC1 presence on the membrane of the lysosomes was monitored by cell fractionation and mTOR expression was determined by immunoblotting.
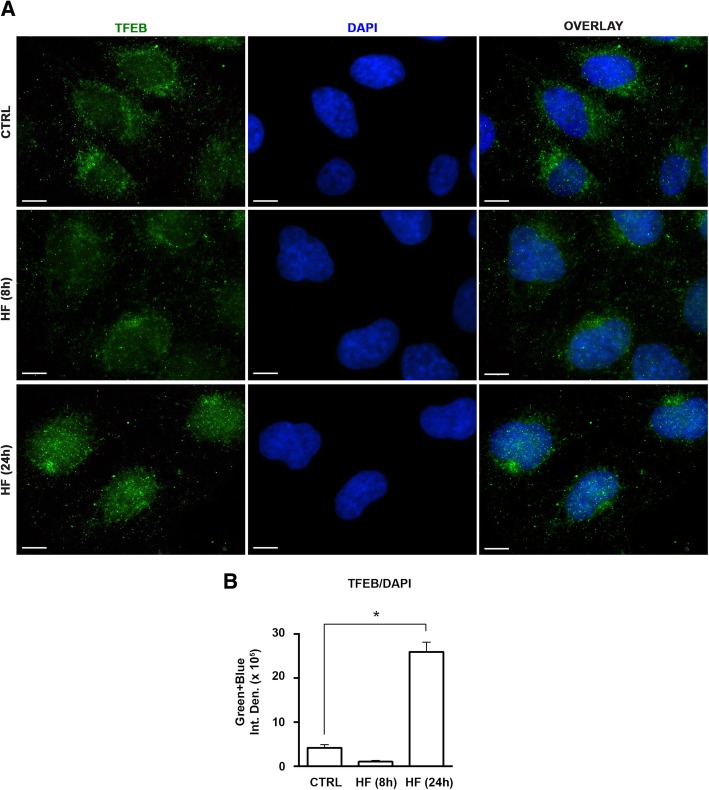
In three different types of human cancer cells (thyroid cancer WRO cells, ovarian cancer OAW-42 cells, and breast cancer MCF-7 cells), HF induced both the AAR and the autophagy pathways time-dependently. In WRO cells, which showed the strongest induction of autophagy and of AAR, global protein synthesis was little if any affected. Consistently, 4E-BP1 and (rp)S6 were phosphorylated. Concomitantly, mTOR expression and activation declined along with its detachment from the lysosomes and its degradation by the proteasome, and with the nuclear translocation of transcription factor EB (TFEB), a transcription factor of many ATG genes. The extra supplementation of proline rescued all these effects.
We demonstrate that the AAR and autophagy are mechanistically linked at the level of mTORC1, and that the lysosome is the central hub of the cross-talk between these two metabolic stress responses.
在氨基酸饥饿的情况下,细胞激活两种主要的保护途径:氨基酸饥饿反应 (AAR),以抑制全局翻译,和自噬,以从冗余自身成分的降解中恢复必需的底物。AAR 和自噬(ATG)是否以及如何相互调节,以及这两条调节途径何时交汇仍然未知。在这里,我们提供了实验证据,证明雷帕霉素 (mTOR) 复合物 1 (mTORC1) 可特异性定位于溶酶体水平,将 AAR 与自噬途径联系起来。
作为 AAR 的诱导剂,我们使用卤夫酮 (HF),一种与脯氨酰-tRNA 合成酶结合的生物碱,从而模拟脯氨酸 (PRO) 的不可用性。通过评估真核翻译起始因子 (eIF) 2α 的磷酸化来确定 AAR 的诱导。通过评估微管相关蛋白 1 轻链 3 同种型 B (LC3B) 和自噬体-1 (p62/SQSTM1) 水平的处理和积累来监测自噬。通过评估 mTOR、(rp)S6 和 4E-BP1 的磷酸化来监测 mTORC1 的活性。通过嘌呤霉素掺入测定法确定总蛋白合成。通过细胞分级分离监测溶酶体膜上 mTORC1 的存在,并通过免疫印迹法确定 mTOR 的表达。
在三种不同类型的人类癌细胞(甲状腺癌细胞 WRO 细胞、卵巢癌细胞 OAW-42 细胞和乳腺癌 MCF-7 细胞)中,HF 时间依赖性地诱导 AAR 和自噬途径。在自噬和 AAR 诱导最强的 WRO 细胞中,总蛋白合成几乎没有受到影响。一致地,4E-BP1 和 (rp)S6 被磷酸化。同时,mTOR 的表达和激活下降,伴随着其从溶酶体上的分离和被蛋白酶体降解,以及转录因子 EB (TFEB) 的核易位,TFEB 是许多 ATG 基因的转录因子。脯氨酸的额外补充挽救了所有这些效应。
我们证明 AAR 和自噬在 mTORC1 水平上是机械连接的,而溶酶体是这两种代谢应激反应之间相互作用的中心枢纽。