Institute for Medical Engineering and Science, and Department of Physics, Massachusetts Institute of Technology, Cambridge, MA, USA.
Department of Medical Biology, Medical University-Plovdiv, Plovdiv, Bulgaria.
Nature. 2019 Jun;570(7761):395-399. doi: 10.1038/s41586-019-1275-3. Epub 2019 Jun 5.
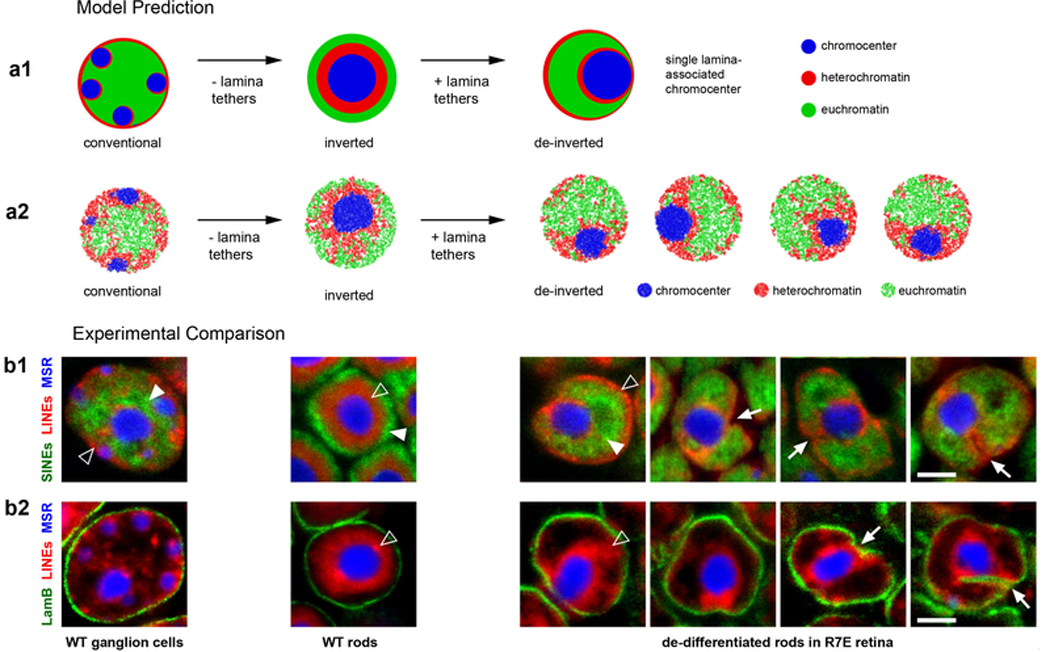
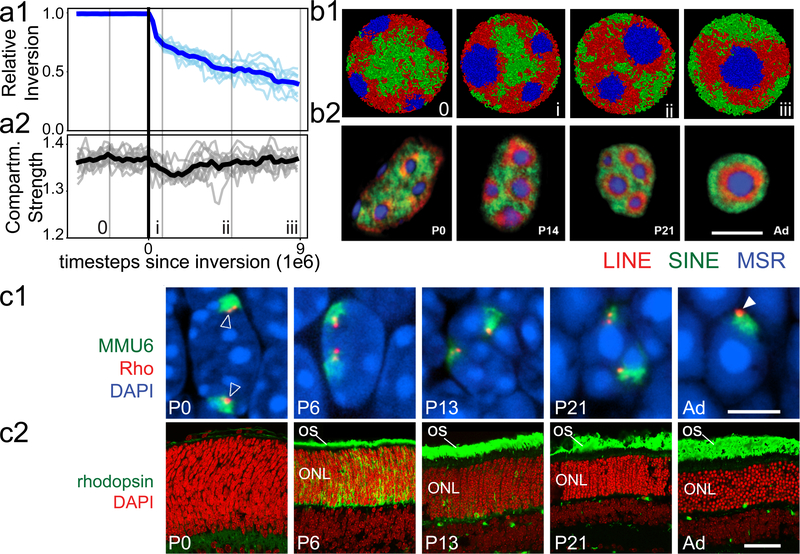
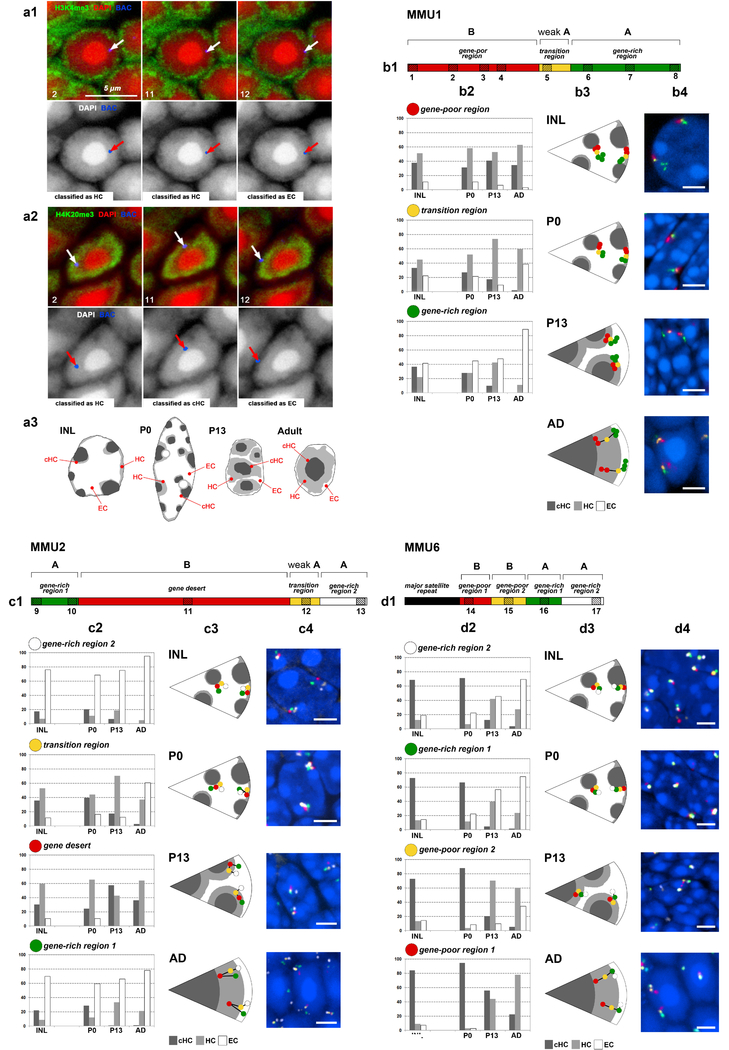
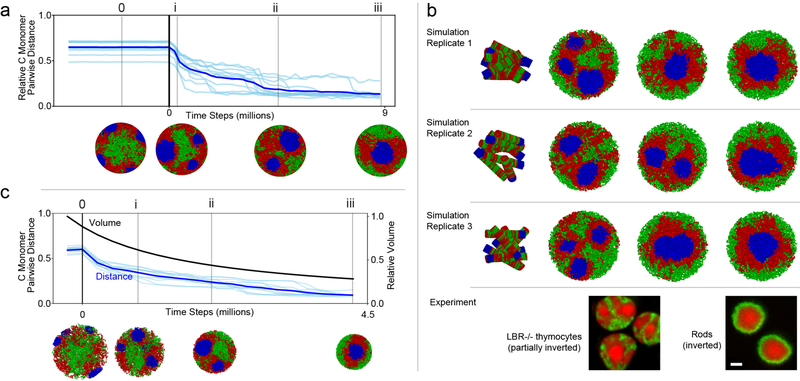
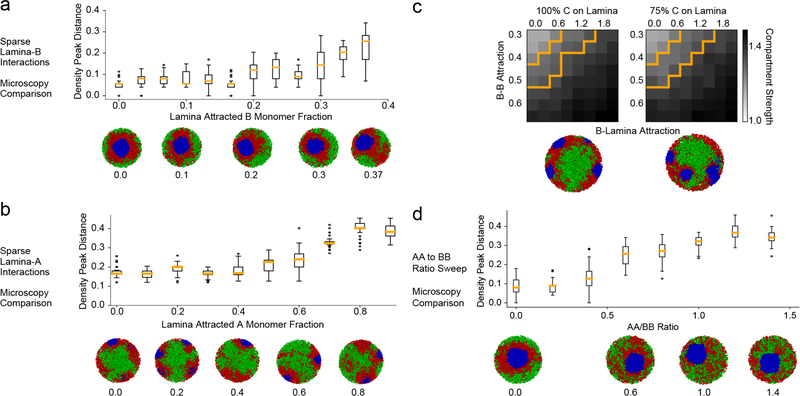
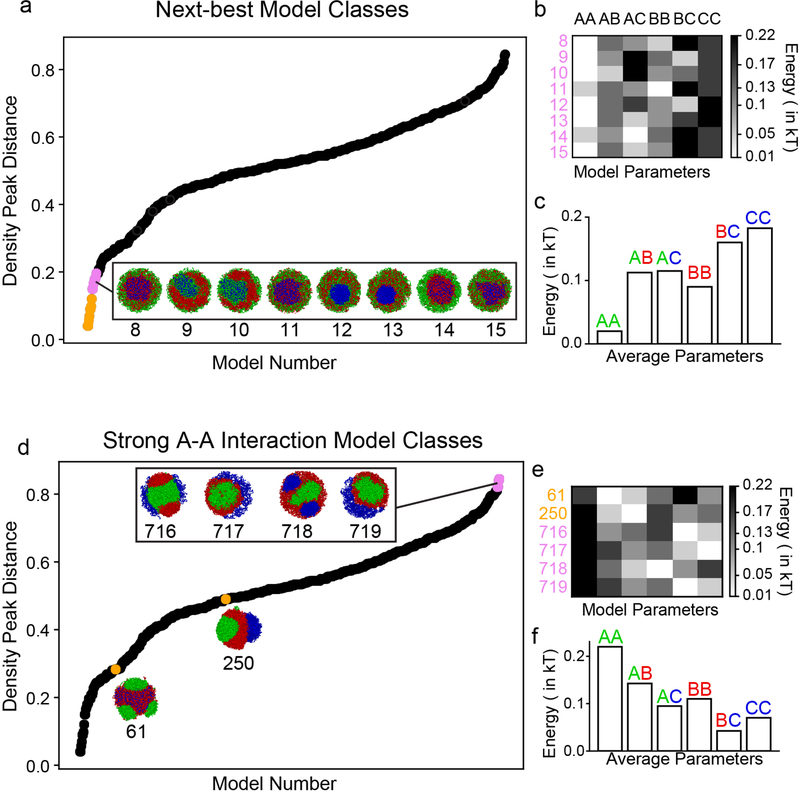
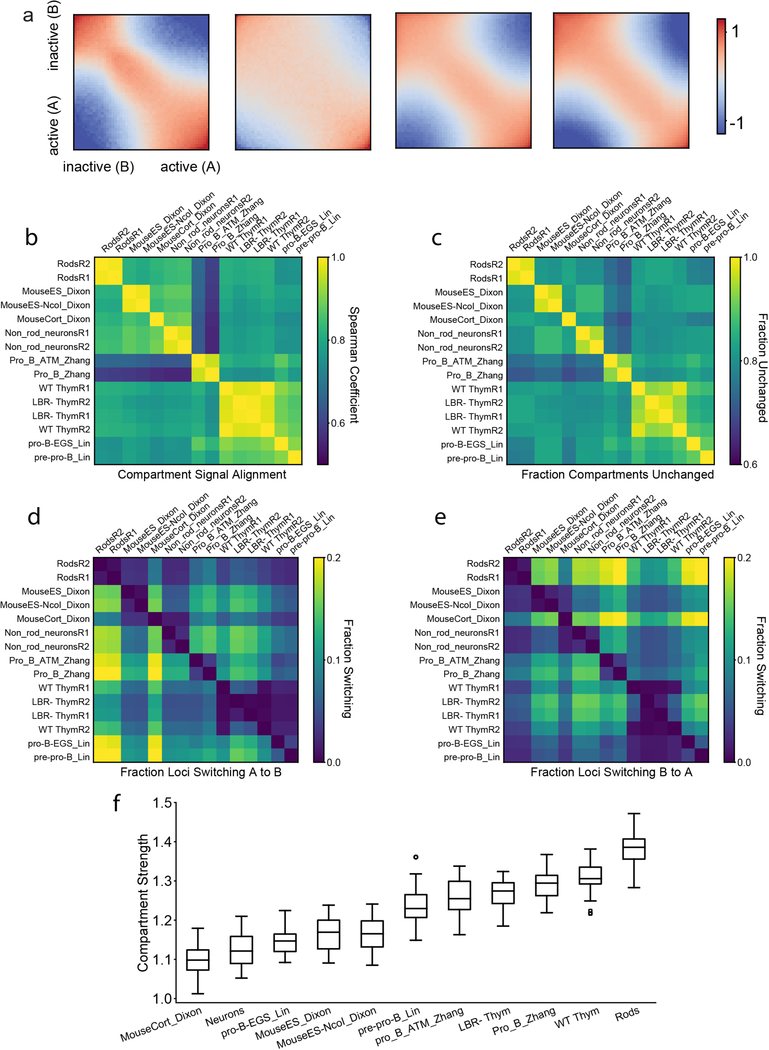
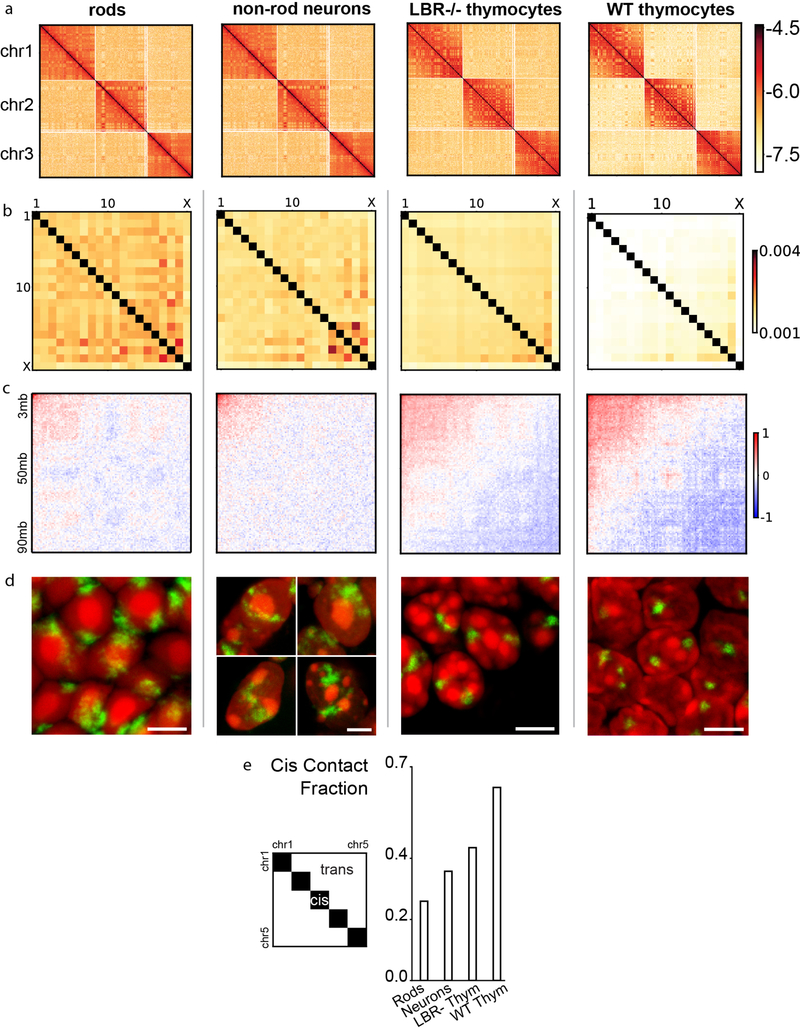
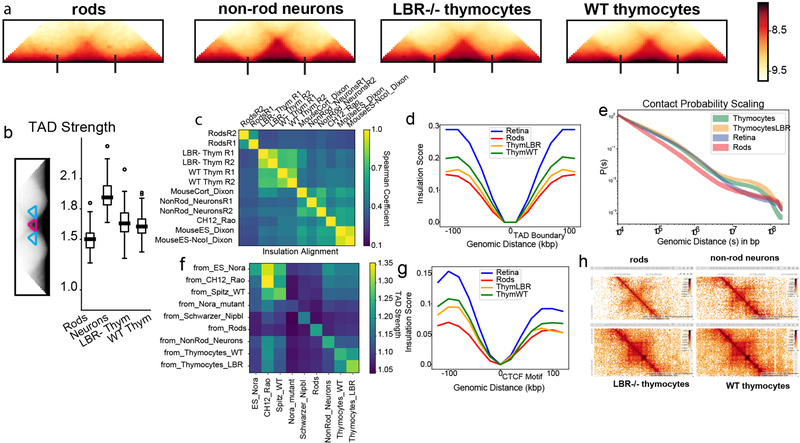
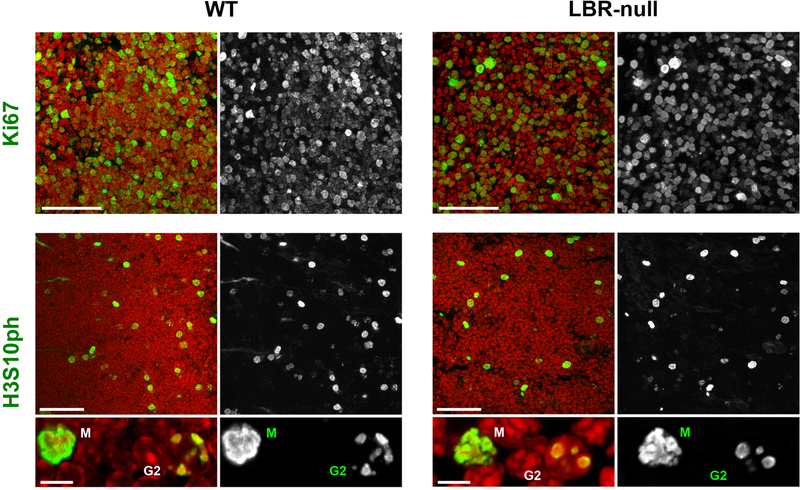
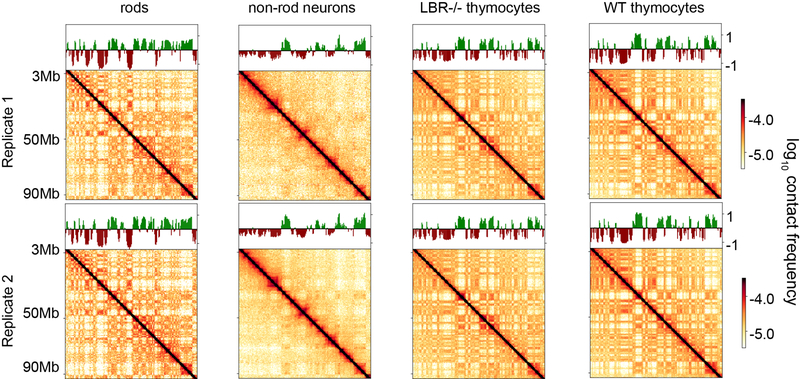
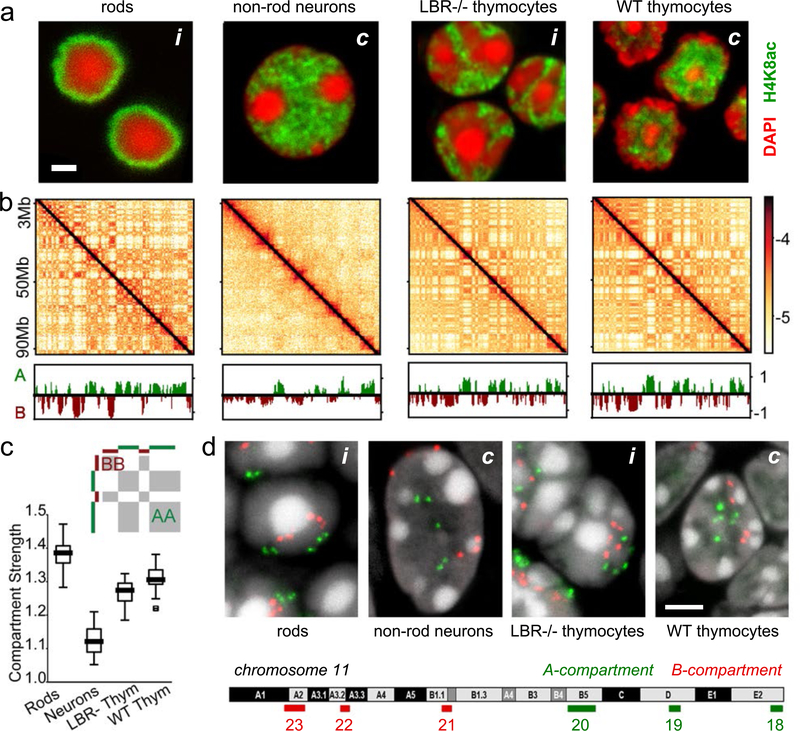
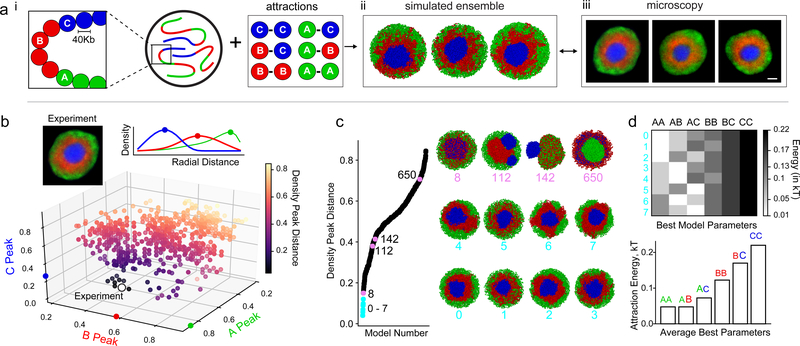
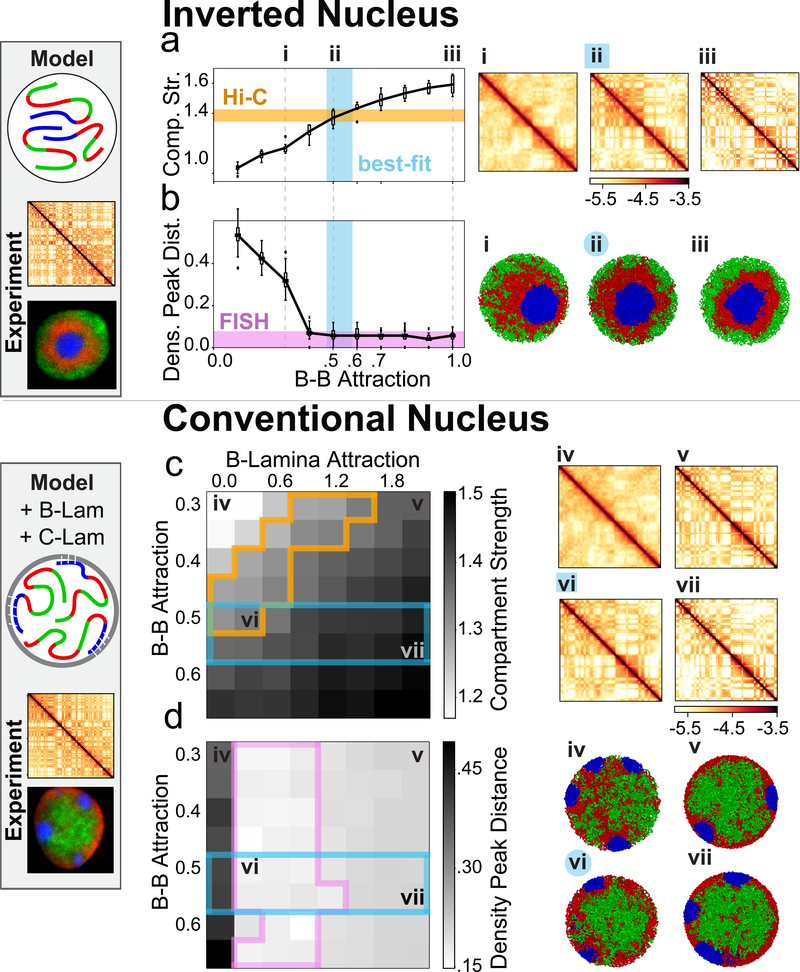
The nucleus of mammalian cells displays a distinct spatial segregation of active euchromatic and inactive heterochromatic regions of the genome. In conventional nuclei, microscopy shows that euchromatin is localized in the nuclear interior and heterochromatin at the nuclear periphery. Genome-wide chromosome conformation capture (Hi-C) analyses show this segregation as a plaid pattern of contact enrichment within euchromatin and heterochromatin compartments, and depletion between them. Many mechanisms for the formation of compartments have been proposed, such as attraction of heterochromatin to the nuclear lamina, preferential attraction of similar chromatin to each other, higher levels of chromatin mobility in active chromatin and transcription-related clustering of euchromatin. However, these hypotheses have remained inconclusive, owing to the difficulty of disentangling intra-chromatin and chromatin-lamina interactions in conventional nuclei. The marked reorganization of interphase chromosomes in the inverted nuclei of rods in nocturnal mammals provides an opportunity to elucidate the mechanisms that underlie spatial compartmentalization. Here we combine Hi-C analysis of inverted rod nuclei with microscopy and polymer simulations. We find that attractions between heterochromatic regions are crucial for establishing both compartmentalization and the concentric shells of pericentromeric heterochromatin, facultative heterochromatin and euchromatin in the inverted nucleus. When interactions between heterochromatin and the lamina are added, the same model recreates the conventional nuclear organization. In addition, our models allow us to rule out mechanisms of compartmentalization that involve strong euchromatin interactions. Together, our experiments and modelling suggest that attractions between heterochromatic regions are essential for the phase separation of the active and inactive genome in inverted and conventional nuclei, whereas interactions of the chromatin with the lamina are necessary to build the conventional architecture from these segregated phases.
哺乳动物细胞的核显示出基因组的活性常染色质和非活性异染色质区域的明显空间分离。在常规核中,显微镜观察显示常染色质定位于核内部,异染色质位于核边缘。全基因组染色体构象捕获(Hi-C)分析显示,这种分离表现为常染色质和异染色质隔室之间的接触富集的棋盘图案,以及它们之间的耗尽。已经提出了许多形成隔室的机制,例如异染色质向核层的吸引、相似染色质彼此的优先吸引、活性染色质中染色质流动性的增加以及常染色质与转录相关的聚集。然而,由于难以分离常规核中的染色质内相互作用和染色质-层相互作用,这些假设仍然没有定论。在夜间哺乳动物的杆状倒置核中,间期染色体的明显重排为阐明空间分隔的机制提供了机会。在这里,我们将倒置杆状核的 Hi-C 分析与显微镜和聚合物模拟相结合。我们发现,异染色质区域之间的吸引力对于建立分隔以及倒置核中着丝粒异染色质、兼性异染色质和常染色质的同心壳是至关重要的。当加入异染色质与层之间的相互作用时,相同的模型再现了常规的核组织。此外,我们的模型使我们能够排除涉及强常染色质相互作用的分隔机制。总之,我们的实验和建模表明,异染色质区域之间的吸引力对于倒置和常规核中活性和非活性基因组的相分离是必不可少的,而染色质与层的相互作用对于从这些分隔的相构建常规结构是必要的。