Degenerative Diseases Program, SBP Medical Discovery Institute, La Jolla, United States.
Molecular Biology and Biochemistry, Gottfried Schatz Research Center, Medical University of Graz, Graz, Austria.
Elife. 2019 Sep 9;8:e49682. doi: 10.7554/eLife.49682.
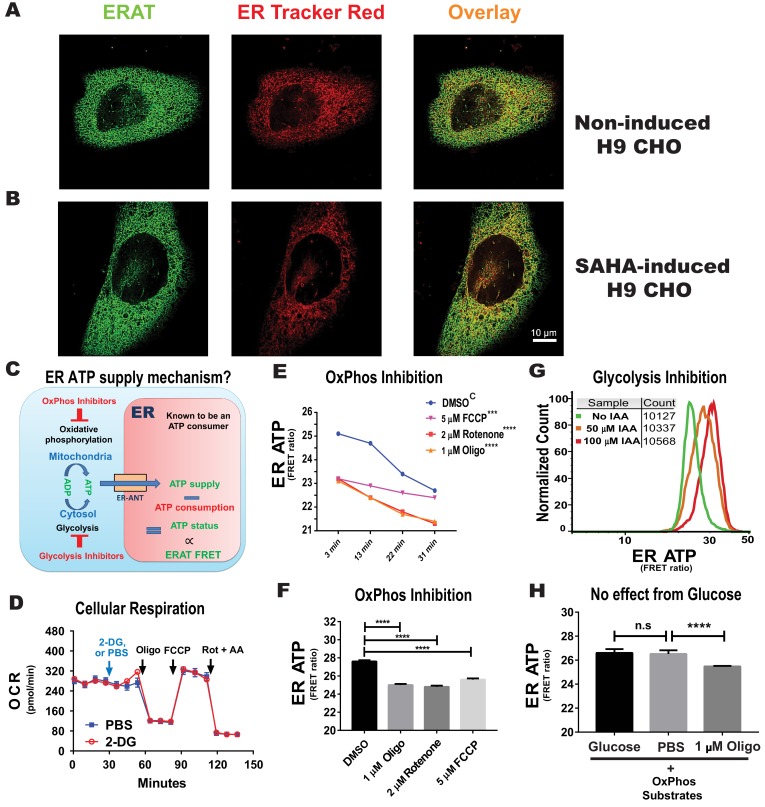
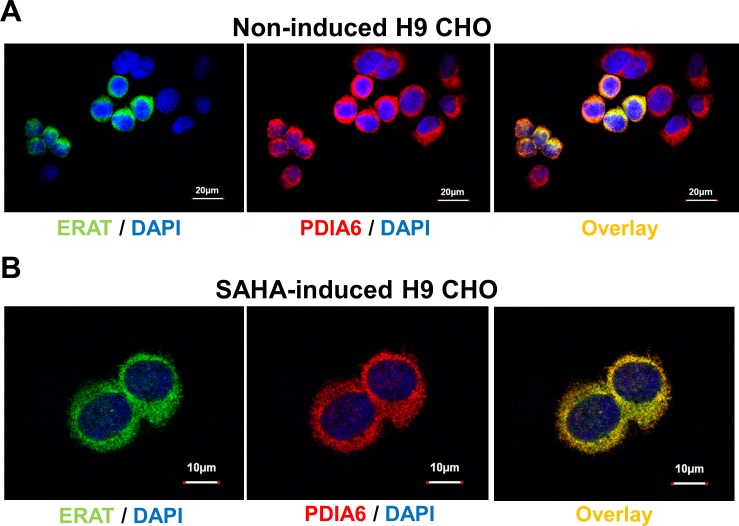
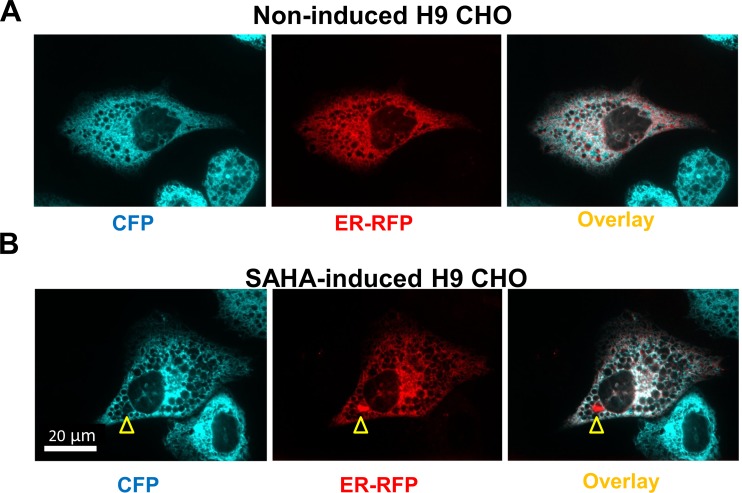
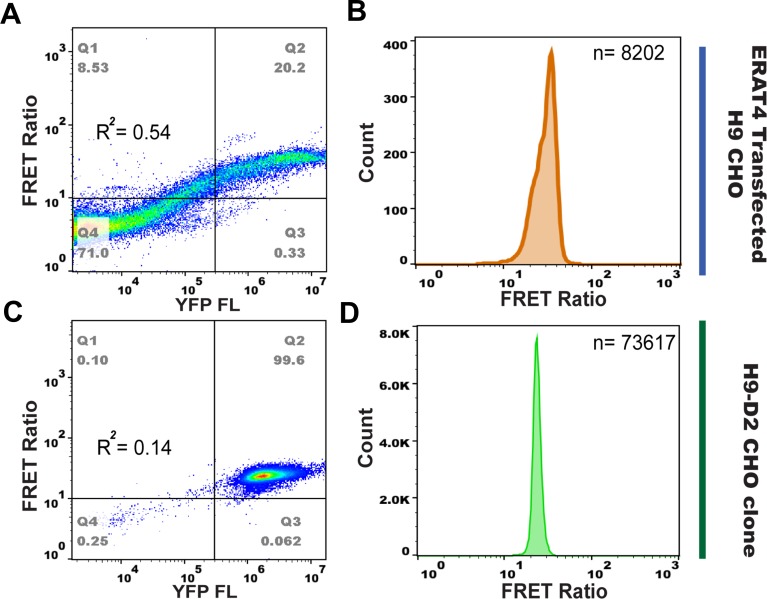
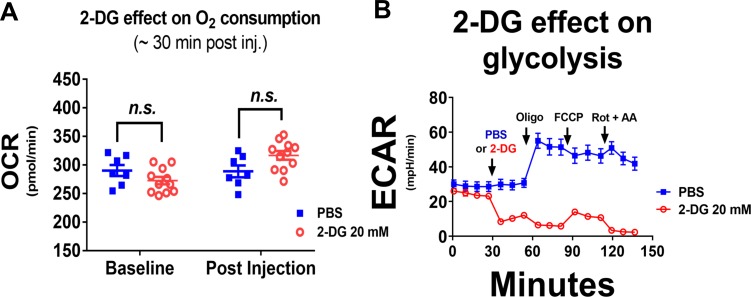
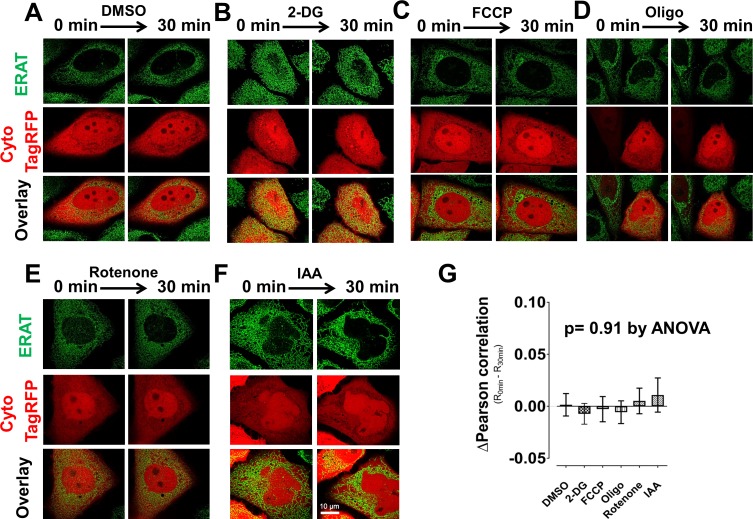
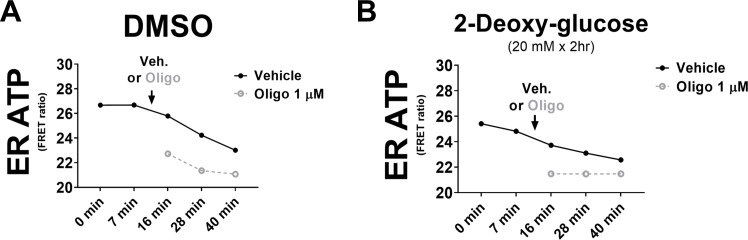
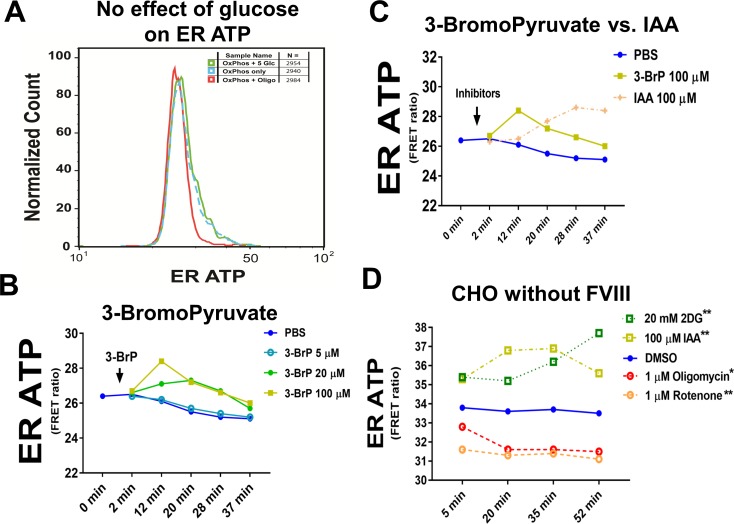
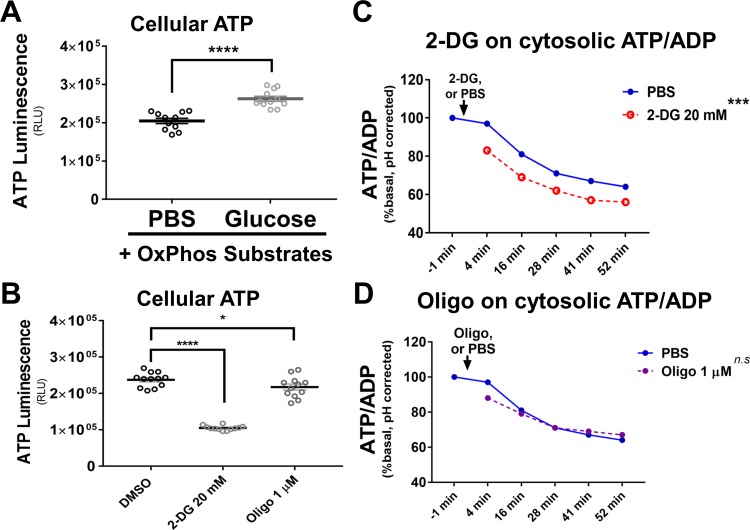
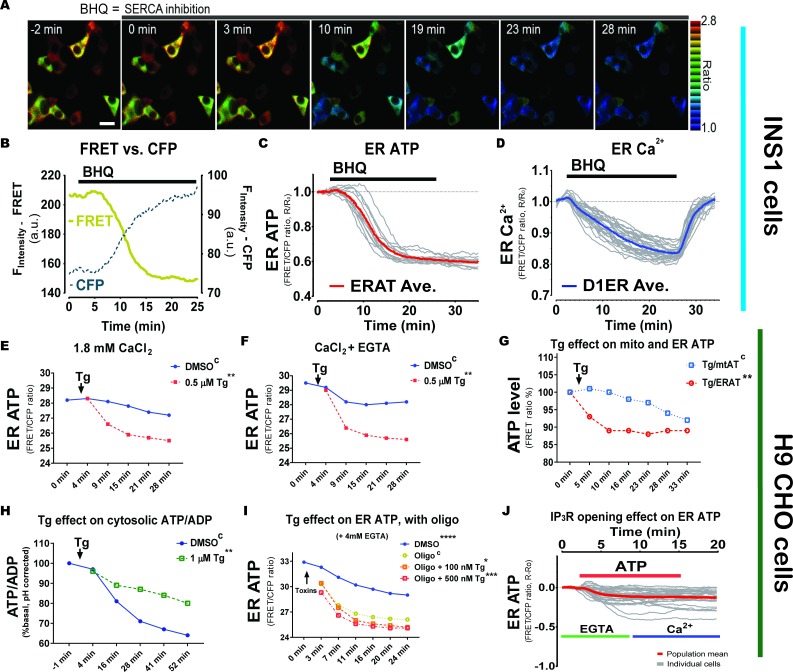
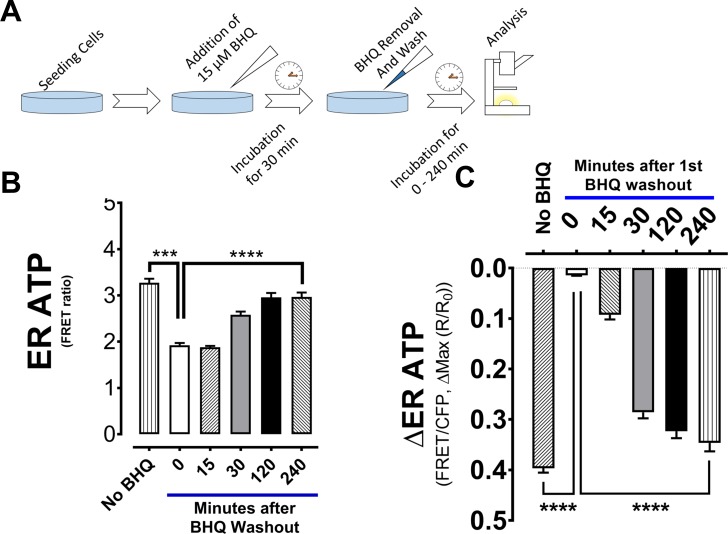
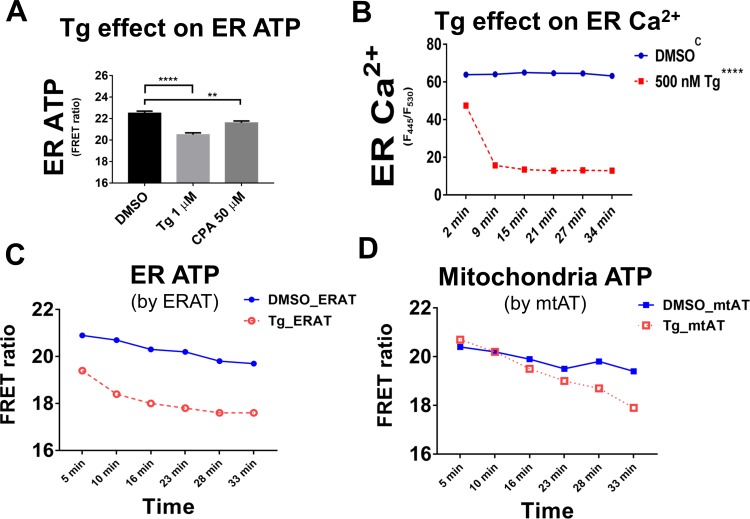
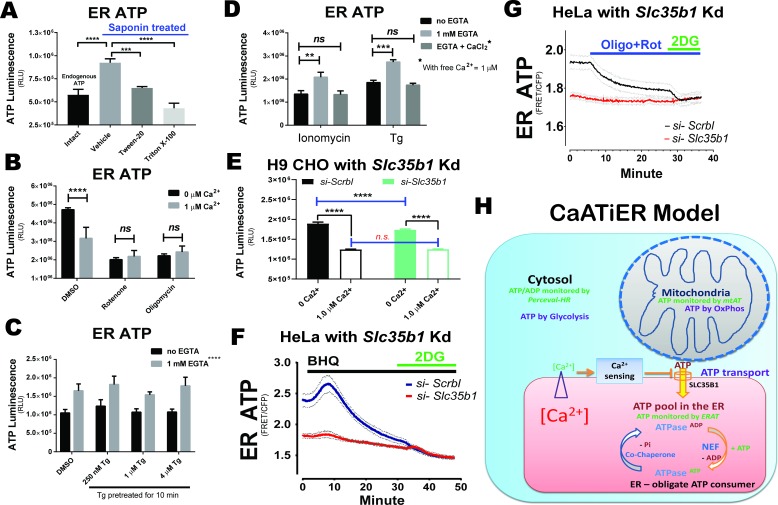
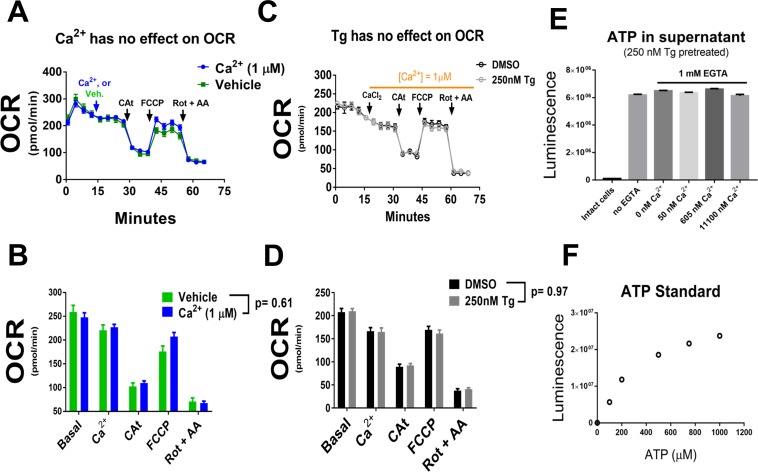
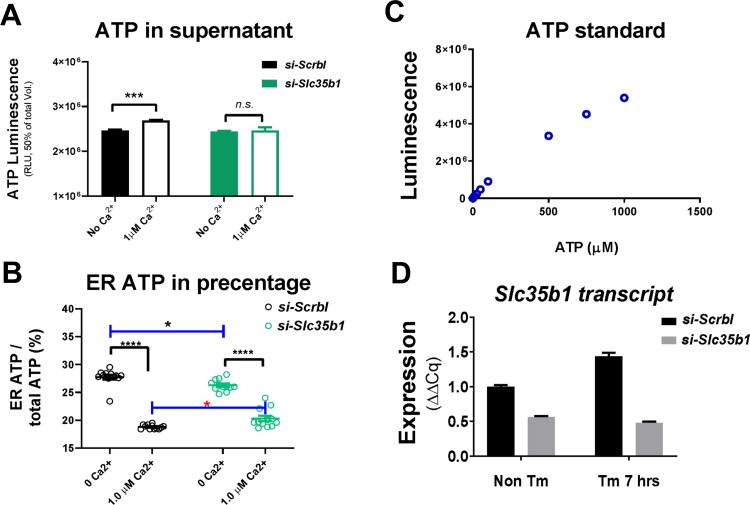
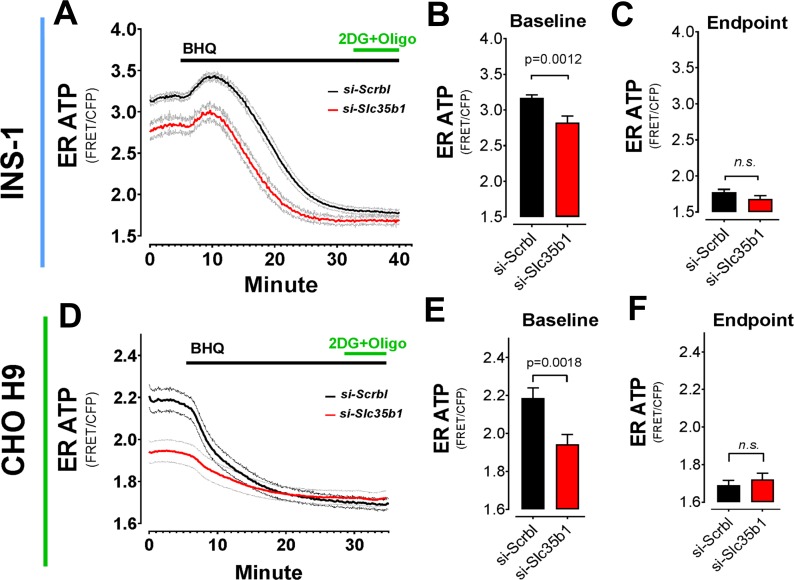
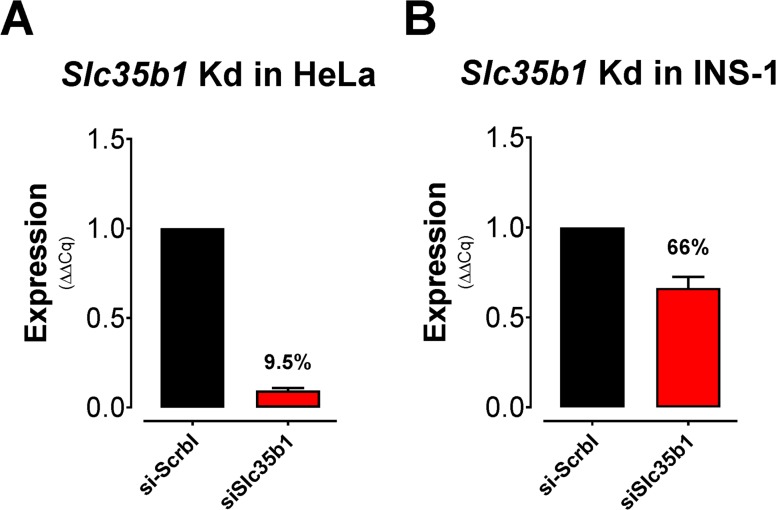
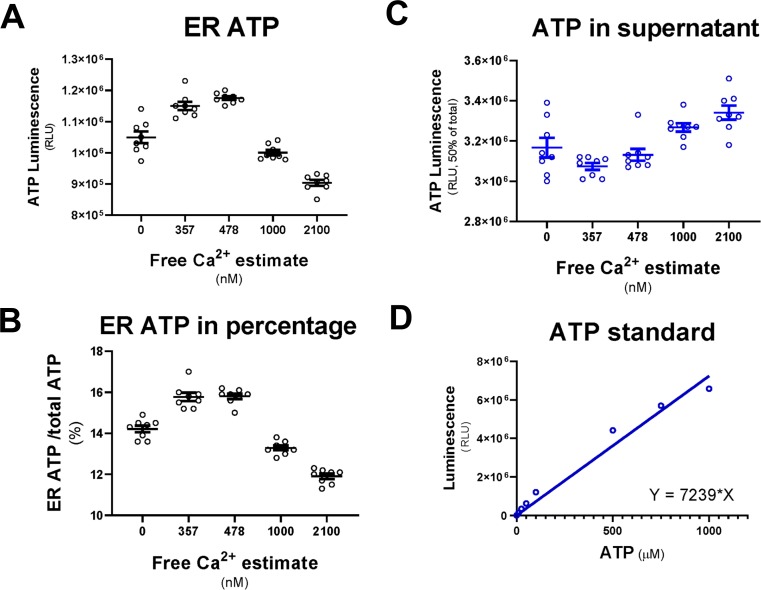
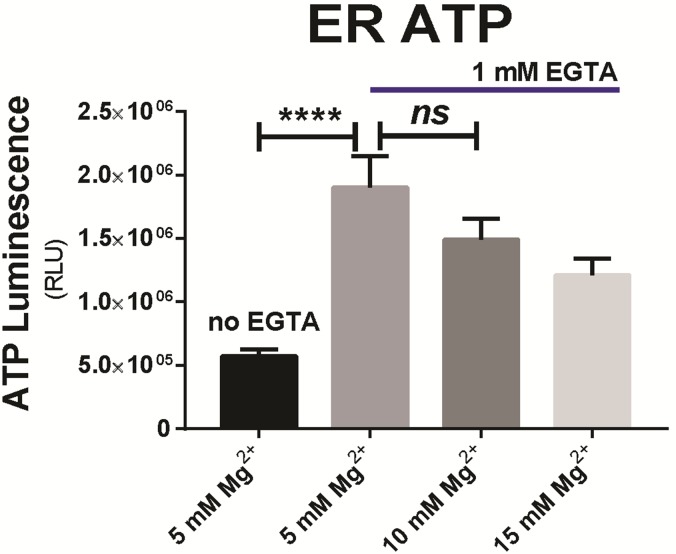
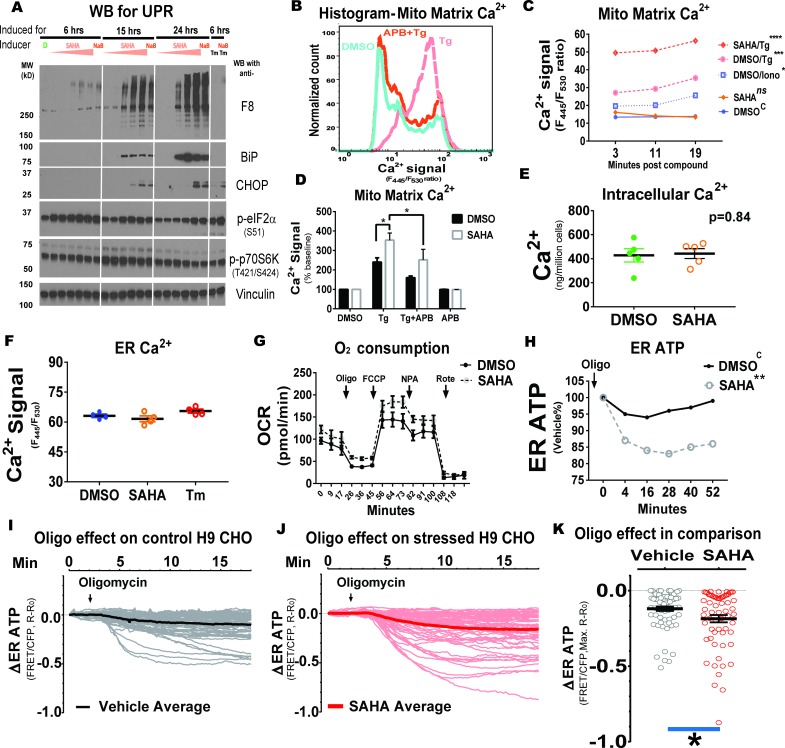
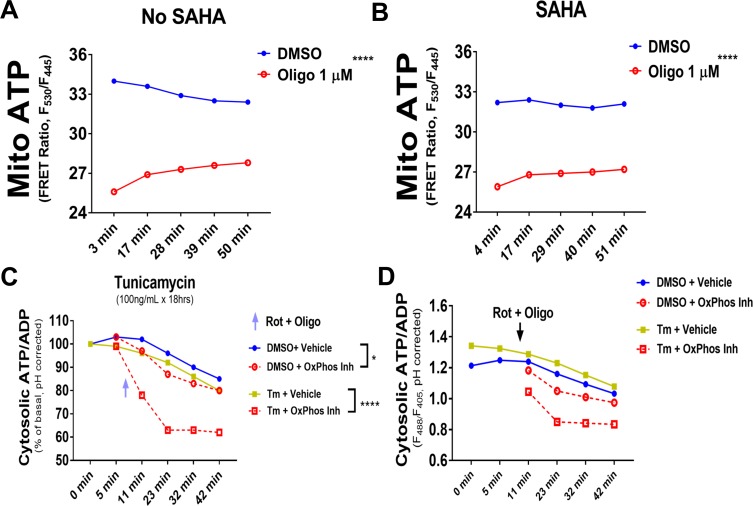
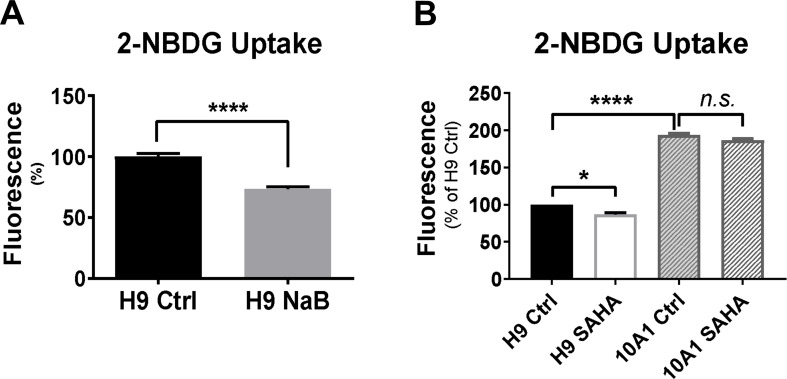
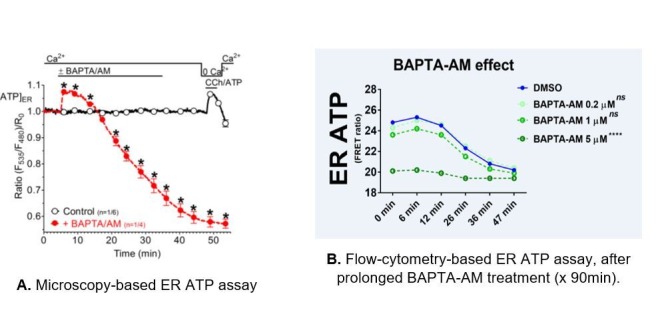
The endoplasmic reticulum () imports ATP and uses energy from ATP hydrolysis for protein folding and trafficking. However, little is known about how this vital ATP transport occurs across the ER membrane. Here, using three commonly used cell lines (CHO, INS1 and HeLa), we report that ATP enters the ER lumen through a cytosolic Ca-antagonized mechanism, or (-ntagonized ransport nto ). Significantly, we show that mitochondria supply ATP to the ER and a SERCA-dependent Ca gradient across the ER membrane is necessary for ATP transport into the ER, through SLC35B1/AXER. We propose that under physiological conditions, increases in cytosolic Ca inhibit ATP import into the ER lumen to limit ER ATP consumption. Furthermore, the ATP level in the ER is readily depleted by oxidative phosphorylation () inhibitors and that ER protein misfolding increases ATP uptake from mitochondria into the ER. These findings suggest that ATP usage in the ER may increase mitochondrial OxPhos while decreasing glycolysis, i.e. an '' effect.
内质网 () 通过利用 ATP 水解产生的能量来输入 ATP,并进行蛋白质折叠和运输。然而,人们对于这种至关重要的 ATP 跨内质网膜运输的机制知之甚少。在这里,我们使用三种常用的细胞系(CHO、INS1 和 HeLa),报告了 ATP 通过细胞质 Ca 拮抗剂机制或 (-ntagonized ransport nto ) 进入内质网腔。重要的是,我们表明线粒体为内质网提供 ATP,并且内质网膜上的 SERCA 依赖性 Ca 梯度对于通过 SLC35B1/AXER 将 ATP 转运到内质网中是必需的。我们提出,在生理条件下,细胞质 Ca 的增加会抑制 ATP 向内质网腔的输入,以限制内质网中 ATP 的消耗。此外,氧化磷酸化 () 抑制剂会迅速耗尽内质网中的 ATP,并且内质网中蛋白质的错误折叠会增加线粒体向内质网摄取 ATP。这些发现表明,内质网中 ATP 的使用可能会增加线粒体的 OxPhos,同时减少糖酵解,即“效”应。