Department of Immunology, St. Jude Children's Research Hospital, Memphis, TN, USA.
Cancer Research UK Cambridge Institute, Li Ka Shing Centre, Cambridge, UK.
Nature. 2019 Sep;573(7775):590-594. doi: 10.1038/s41586-019-1551-2. Epub 2019 Sep 11.
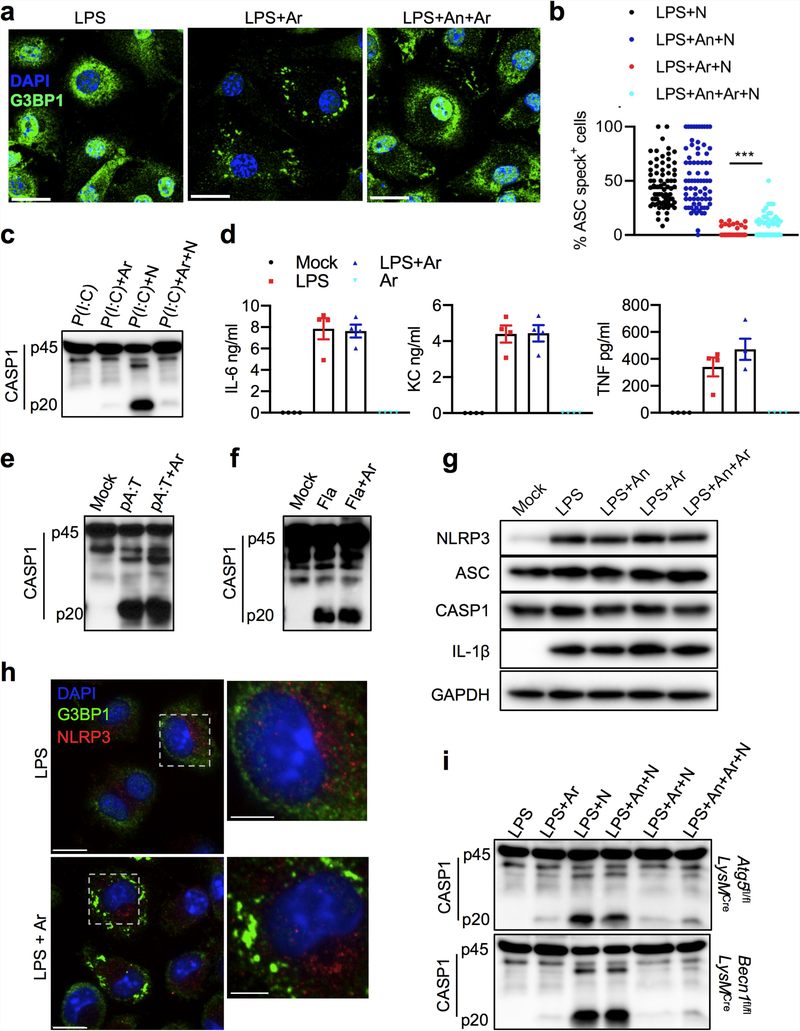
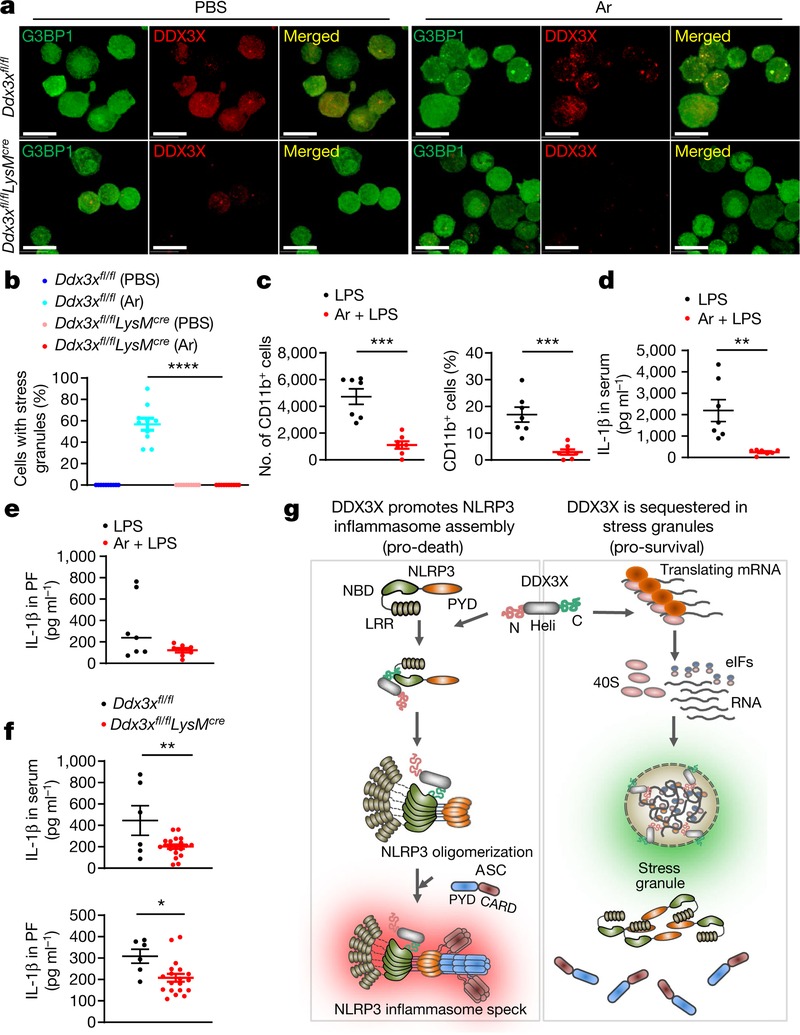
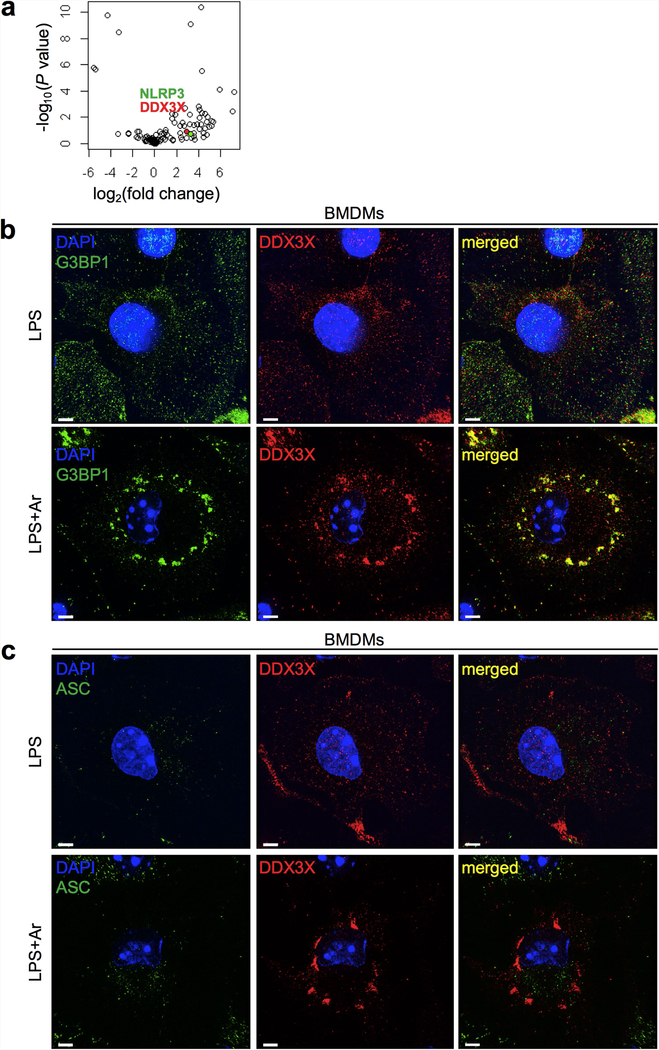
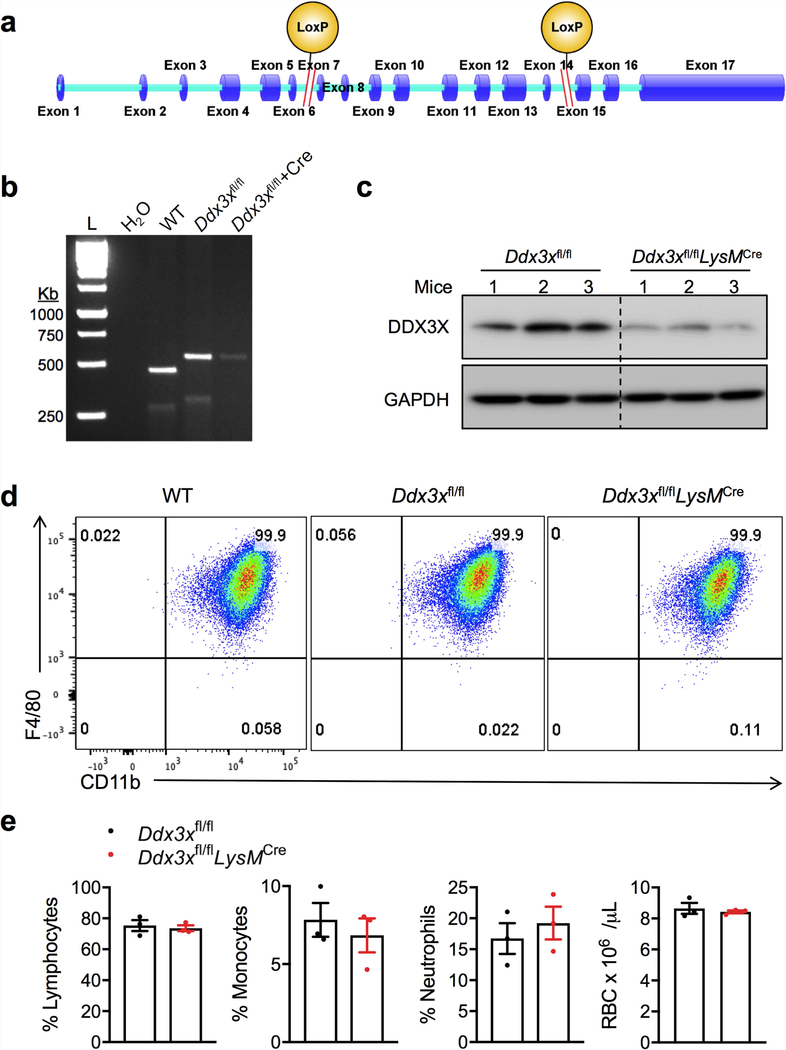
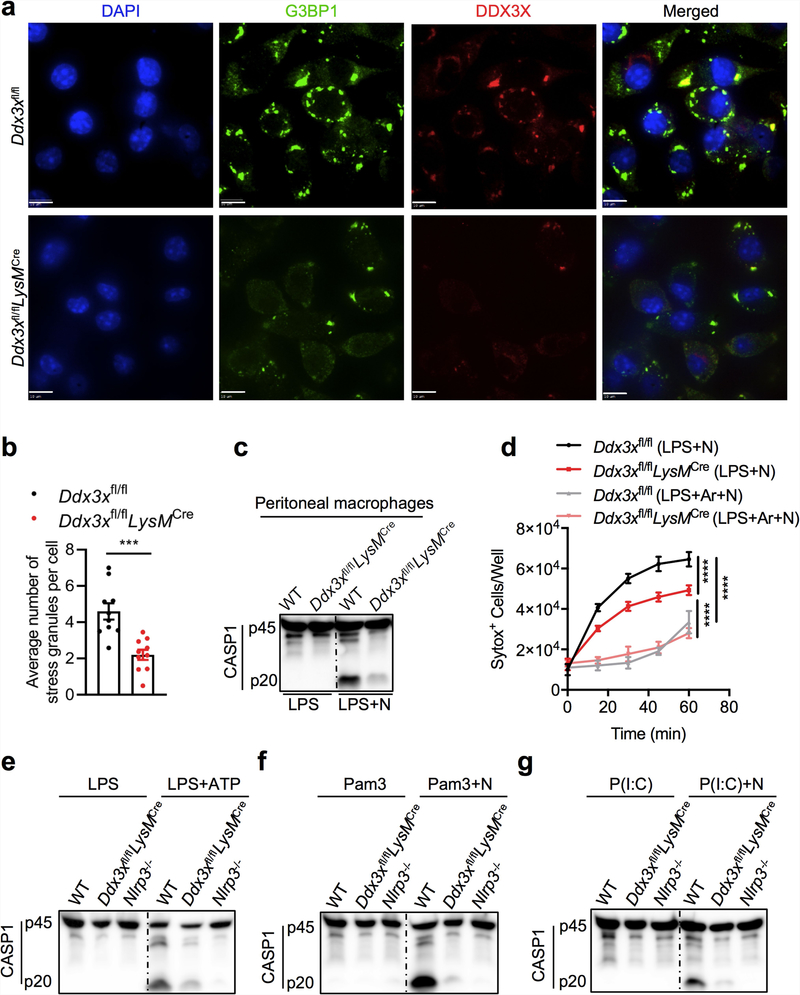
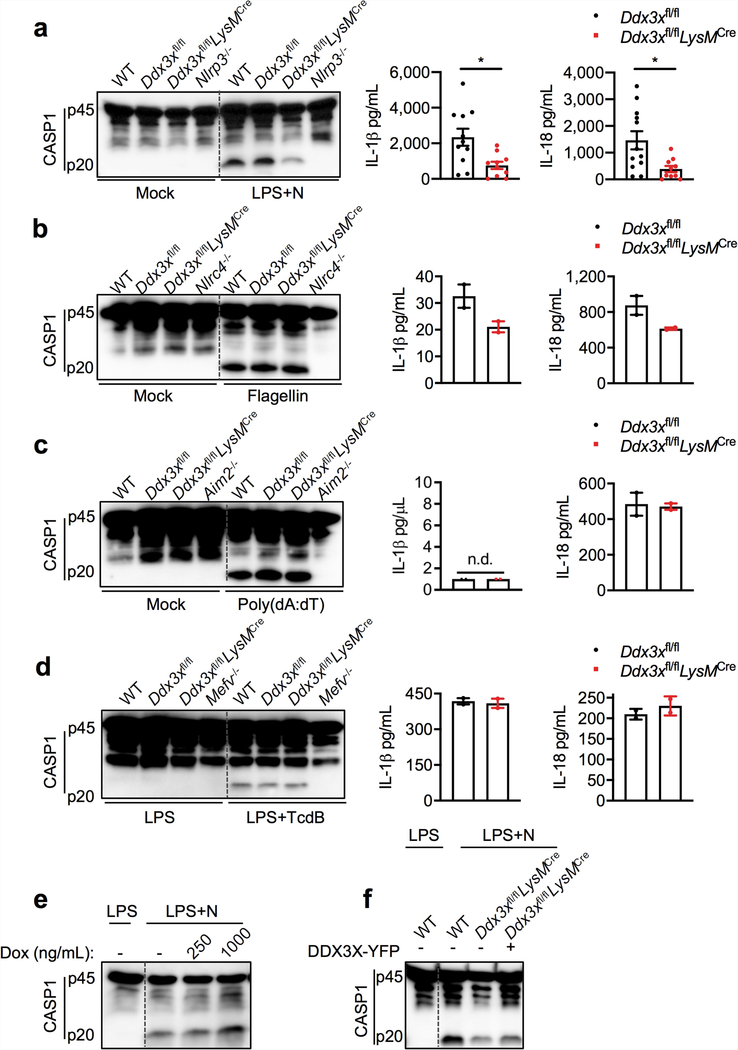
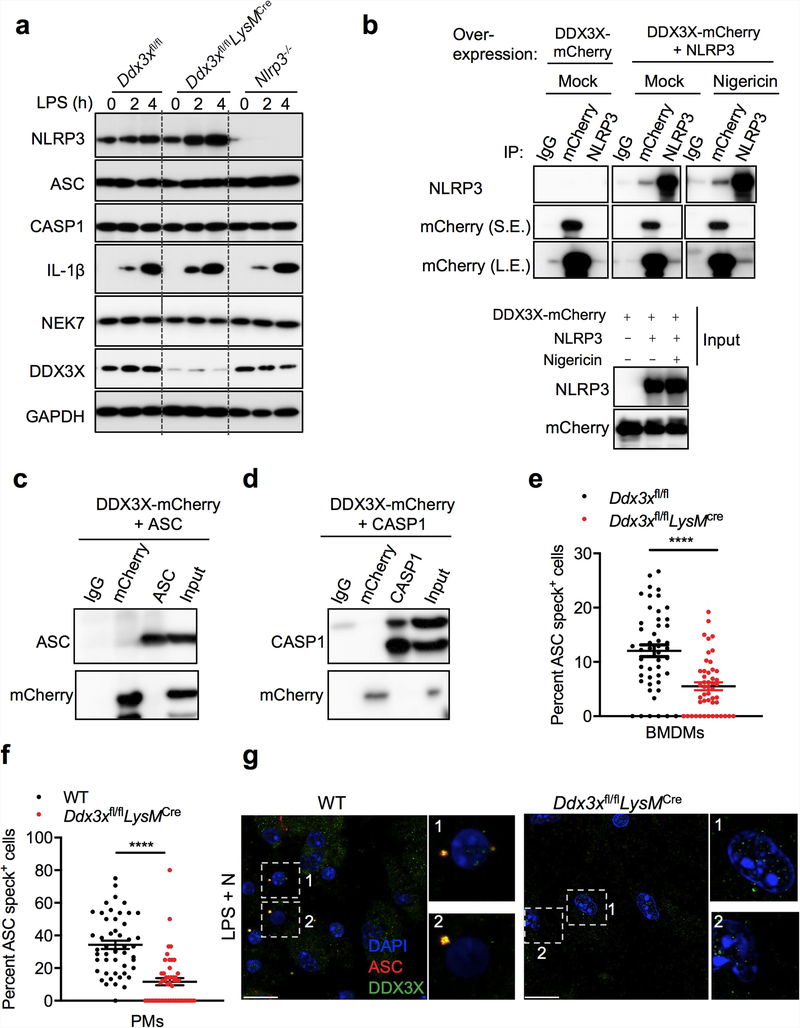
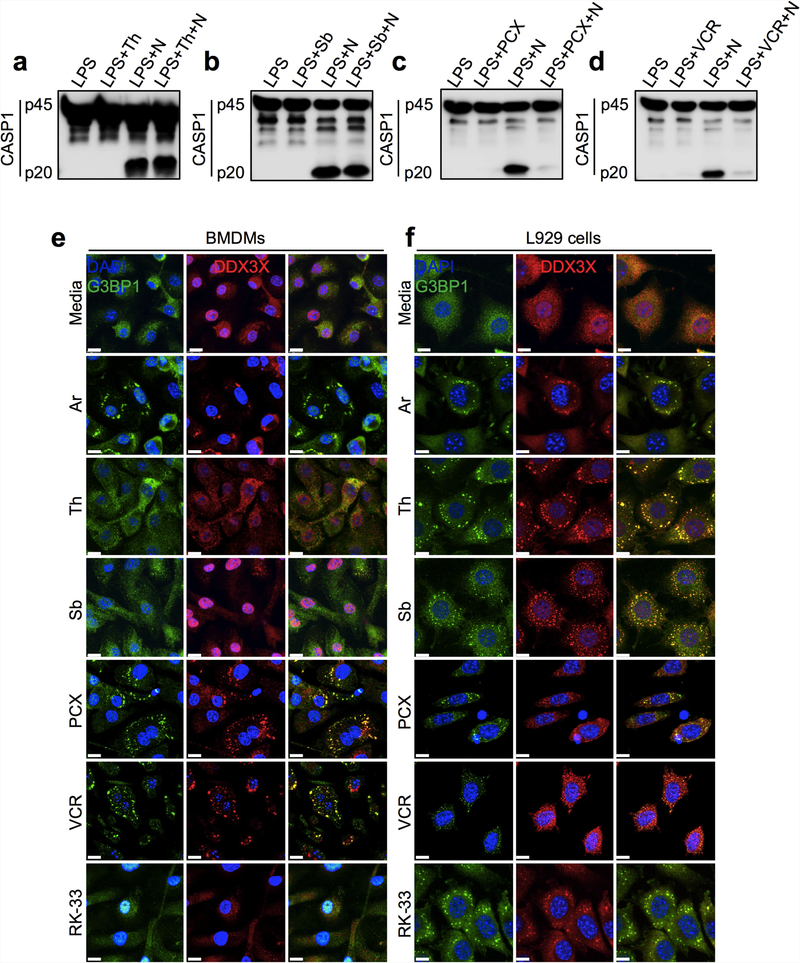
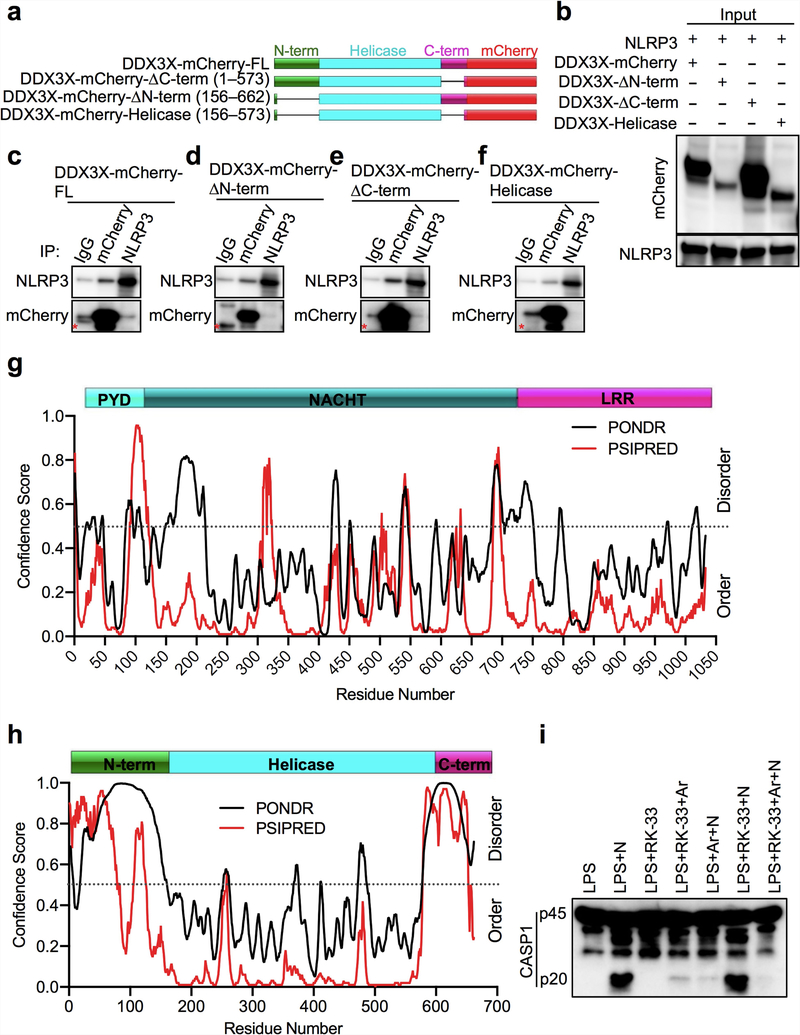
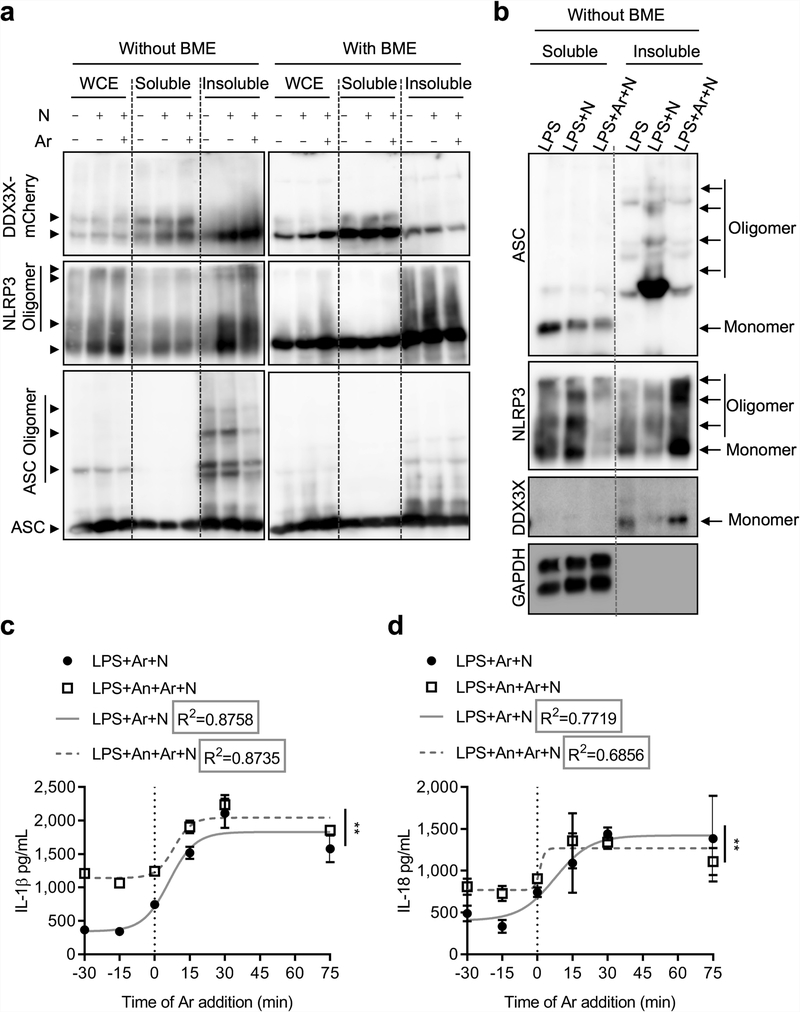
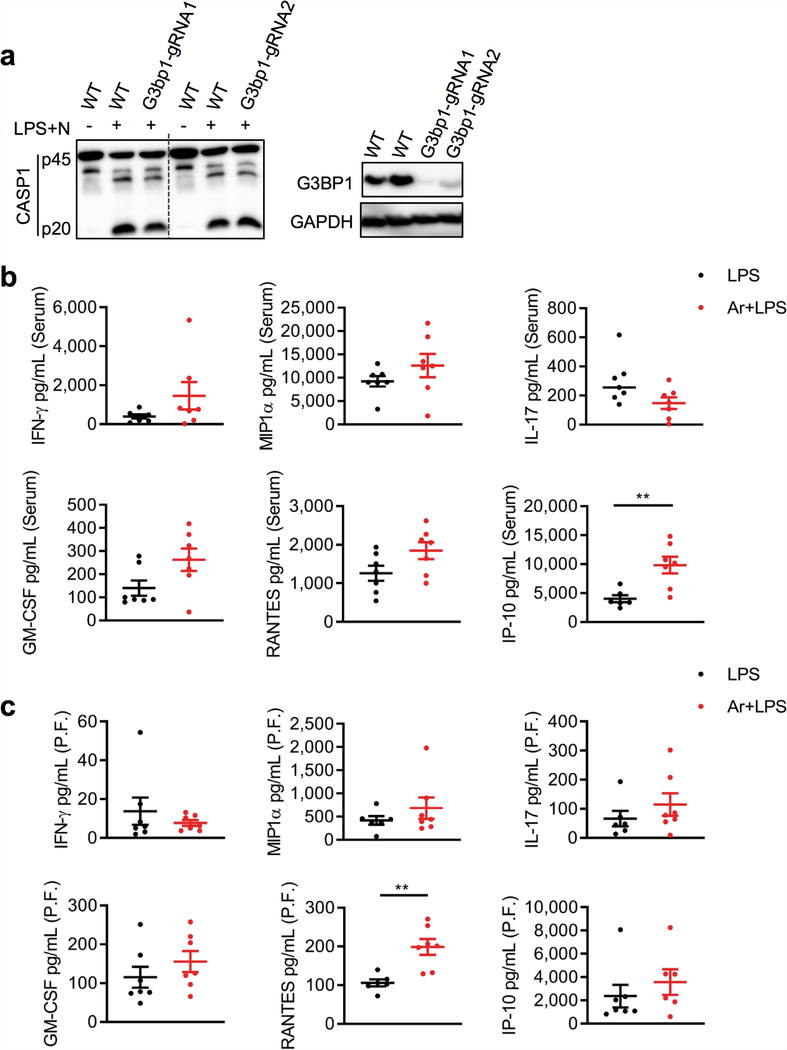
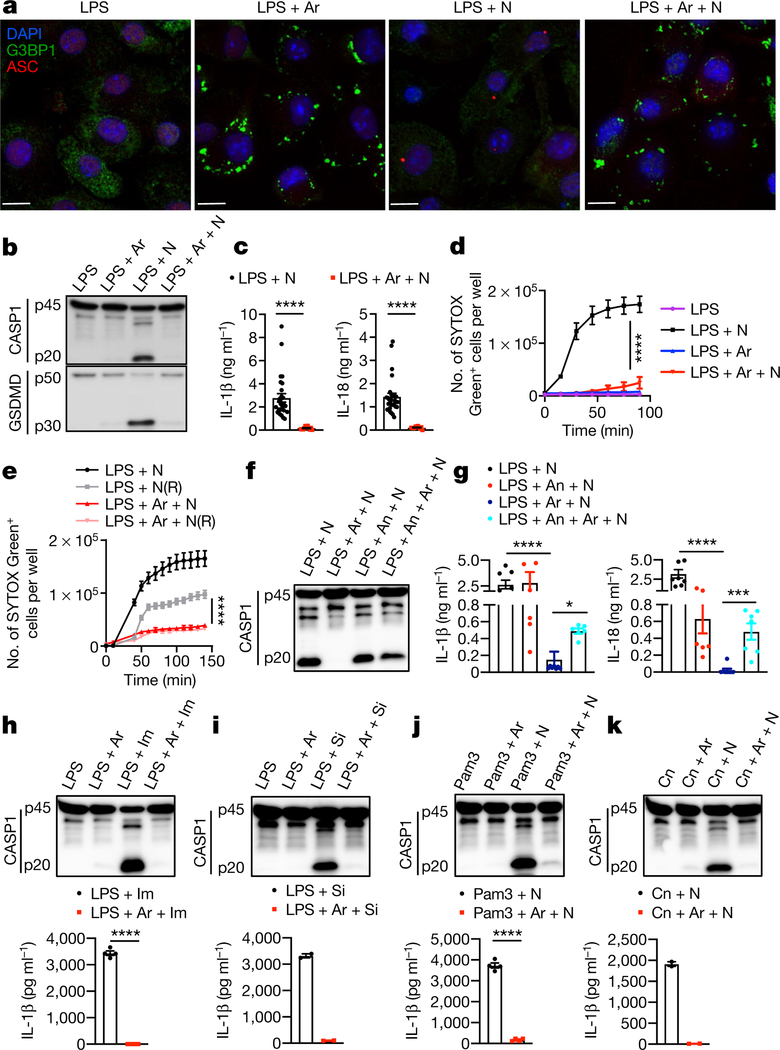
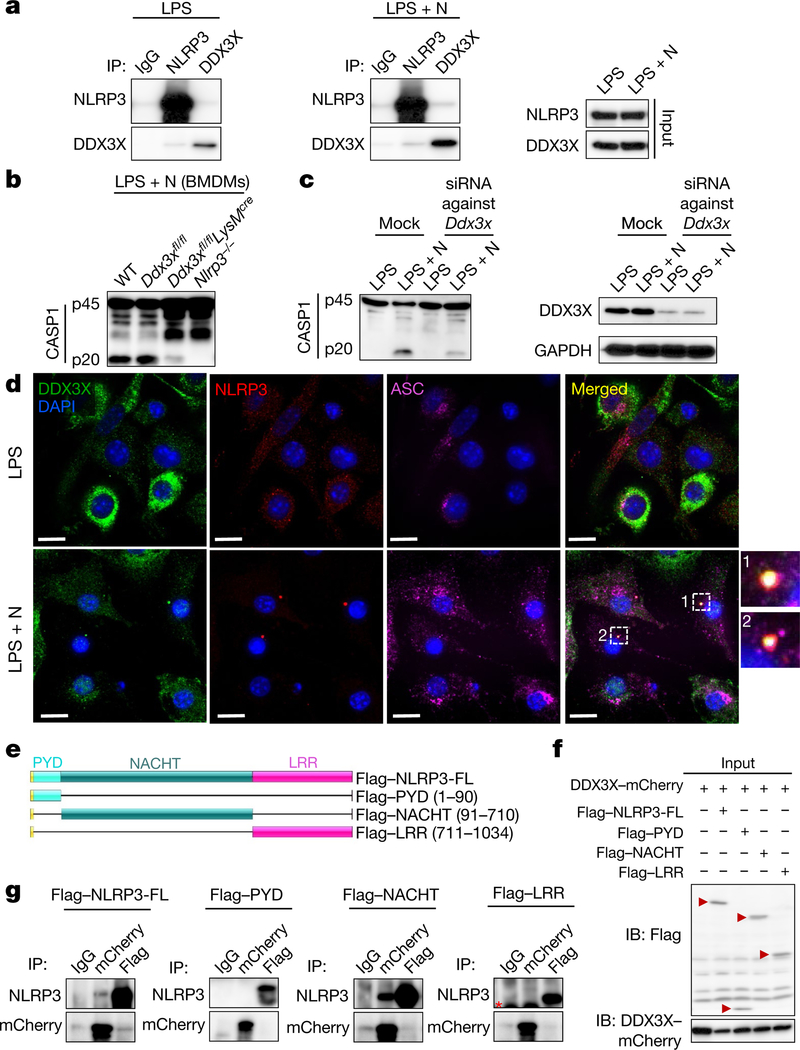
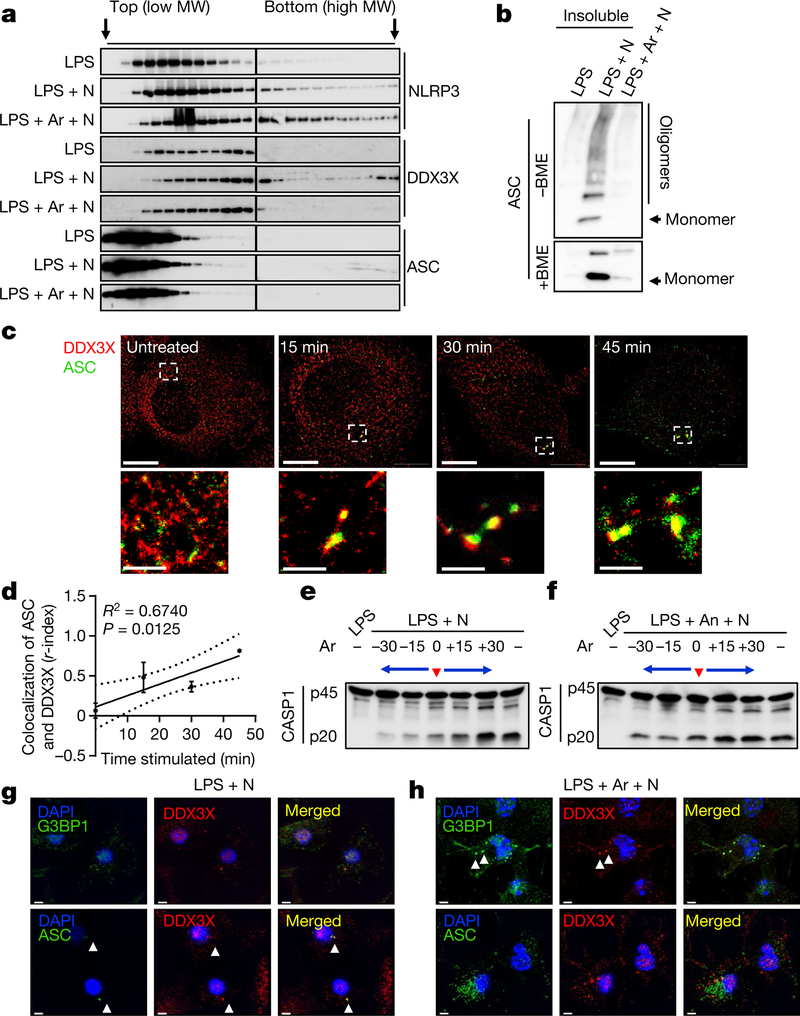
The cellular stress response has a vital role in regulating homeostasis by modulating cell survival and death. Stress granules are cytoplasmic compartments that enable cells to survive various stressors. Defects in the assembly and disassembly of stress granules are linked to neurodegenerative diseases, aberrant antiviral responses and cancer. Inflammasomes are multi-protein heteromeric complexes that sense molecular patterns that are associated with damage or intracellular pathogens, and assemble into cytosolic compartments known as ASC specks to facilitate the activation of caspase-1. Activation of inflammasomes induces the secretion of interleukin (IL)-1β and IL-18 and drives cell fate towards pyroptosis-a form of programmed inflammatory cell death that has major roles in health and disease. Although both stress granules and inflammasomes can be triggered by the sensing of cellular stress, they drive contrasting cell-fate decisions. The crosstalk between stress granules and inflammasomes and how this informs cell fate has not been well-studied. Here we show that the induction of stress granules specifically inhibits NLRP3 inflammasome activation, ASC speck formation and pyroptosis. The stress granule protein DDX3X interacts with NLRP3 to drive inflammasome activation. Assembly of stress granules leads to the sequestration of DDX3X, and thereby the inhibition of NLRP3 inflammasome activation. Stress granules and the NLRP3 inflammasome compete for DDX3X molecules to coordinate the activation of innate responses and subsequent cell-fate decisions under stress conditions. Induction of stress granules or loss of DDX3X in the myeloid compartment leads to a decrease in the production of inflammasome-dependent cytokines in vivo. Our findings suggest that macrophages use the availability of DDX3X to interpret stress signals and choose between pro-survival stress granules and pyroptotic ASC specks. Together, our data demonstrate the role of DDX3X in driving NLRP3 inflammasome and stress granule assembly, and suggest a rheostat-like mechanistic paradigm for regulating live-or-die cell-fate decisions under stress conditions.
细胞应激反应通过调节细胞存活和死亡在维持体内平衡方面起着至关重要的作用。应激颗粒是细胞质隔室,使细胞能够耐受各种应激源。应激颗粒的组装和拆卸缺陷与神经退行性疾病、异常抗病毒反应和癌症有关。炎症小体是多蛋白异源复合物,可感知与损伤或细胞内病原体相关的分子模式,并组装成称为 ASC 斑点的细胞溶质隔室,以促进半胱天冬酶-1 的激活。炎症小体的激活诱导白细胞介素 (IL)-1β 和 IL-18 的分泌,并促使细胞命运朝向细胞焦亡——一种程序性炎症细胞死亡形式,在健康和疾病中具有重要作用。尽管应激颗粒和炎症小体都可以通过细胞应激的感应而触发,但它们驱动着相反的细胞命运决定。应激颗粒和炎症小体之间的串扰以及这如何影响细胞命运尚未得到很好的研究。在这里,我们表明应激颗粒的诱导特异性抑制 NLRP3 炎症小体的激活、ASC 斑点的形成和细胞焦亡。应激颗粒蛋白 DDX3X 与 NLRP3 相互作用以驱动炎症小体的激活。应激颗粒的组装导致 DDX3X 的隔离,从而抑制 NLRP3 炎症小体的激活。应激颗粒和 NLRP3 炎症小体竞争 DDX3X 分子,以协调在应激条件下固有反应的激活和随后的细胞命运决定。髓系细胞中应激颗粒的诱导或 DDX3X 的缺失导致体内炎症小体依赖性细胞因子的产生减少。我们的发现表明,巨噬细胞利用 DDX3X 的可用性来解释应激信号,并在生存应激颗粒和细胞焦亡 ASC 斑点之间做出选择。总之,我们的数据表明 DDX3X 在驱动 NLRP3 炎症小体和应激颗粒组装中的作用,并提出了一种在应激条件下调节生与死细胞命运决定的变阻器样机制范例。