Swart Reuben M, le Roux Francois, Naude Andre, de Jongh Nicolaas W, Nicol Willie
Department of Chemical Engineering, University of Pretoria, Lynnwood Road, Hatfield, 0002 Pretoria South Africa.
Biotechnol Biofuels. 2020 Feb 1;13:22. doi: 10.1186/s13068-020-1664-8. eCollection 2020.
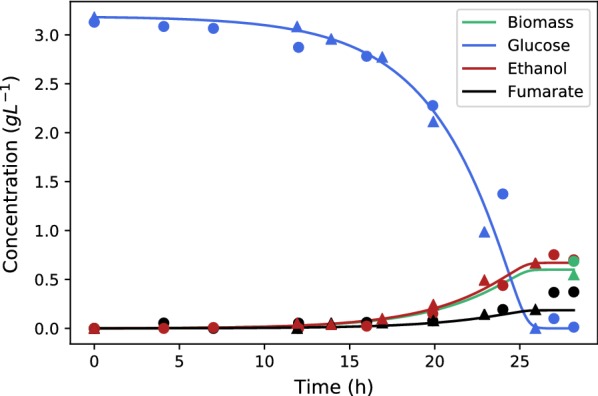
The four-carbon dicarboxylic acids of the tricarboxylic acid cycle (malate, fumarate and succinate) remain promising bio-based alternatives to various precursor chemicals derived from fossil-based feed stocks. The double carbon bond in fumarate, in addition to the two terminal carboxylic groups, opens up an array of downstream reaction possibilities, where replacement options for petrochemical derived maleic anhydride are worth mentioning. To date the most promising organism for producing fumarate is (ATCC 20344, also referred to as ) that naturally excretes fumarate under nitrogen-limited conditions. Fumarate excretion in is always associated with the co-excretion of ethanol, an unwanted metabolic product from the fermentation. Attempts to eliminate ethanol production classically focus on enhanced oxygen availability within the mycelium matrix. In this study our immobilised process was employed to investigate and utilise the Crabtree characteristics of the organism in order to establish the limits of ethanol by-product formation under growth and non-growth conditions.
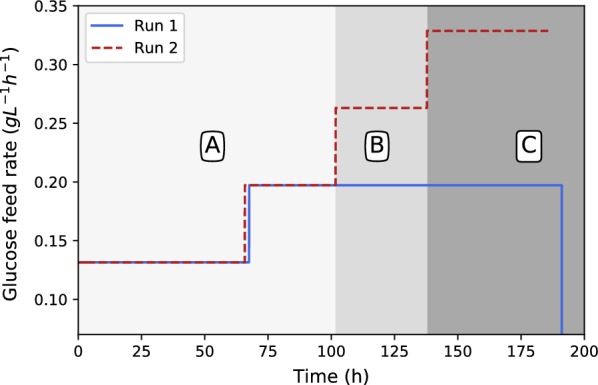
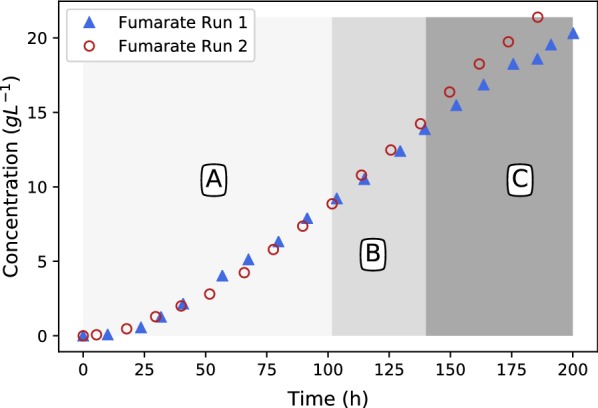
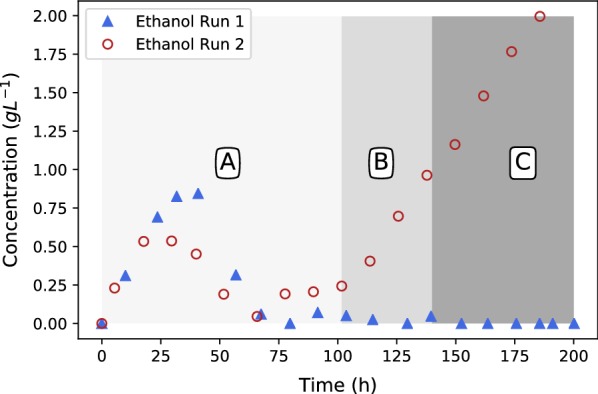
All fermentations were performed with either nitrogen excess (growth phase) or nitrogen limitation (production phase) where medium replacements were done between the growth and the production phase. Initial experiments employed excess glucose for both growth and production, while the oxygen partial pressure was varied between a dissolved oxygen of 18.4% and 85%. Ethanol was formed during both growth and production phases and the oxygen partial pressure had zero influence on the response. Results clearly indicated that possible anaerobic zones within the mycelium were not responsible for ethanol formation, hinting that ethanol is formed under fully aerobic conditions as a metabolic overflow product. For Crabtree-positive organisms like ethanol overflow is manipulated by controlling the glucose input to the fermentation. The same strategy was employed for for both growth and production fermentations. It was shown that all ethanol can be eliminated during growth for a glucose addition rate of . The production phase behaved in a similar manner, where glucose addition of resulted in fumarate production of and a yield of fumarate on glucose. Further investigation into the effect of glucose addition revealed that ethanol overflow commences at a glucose addition rate of on biomass, while the maximum glucose uptake rate was established to be between 0.426 and .
The results conclusively prove that is a Crabtree-positive organism and that the characteristic can be utilised to completely discard ethanol by-product formation. A state referred to as "homofumarate production" was illustrated, where all carbon input exits the cell as either fumarate or respiratory . The highest biomass-based "homofumarate production": rate of achieved a yield of on glucose, indicating the bounds for developing an ethanol free process. The control strategy employed in this study in conjunction with the uncomplicated scalability of the immobilised process provides new direction for further developing bio-fumarate production.
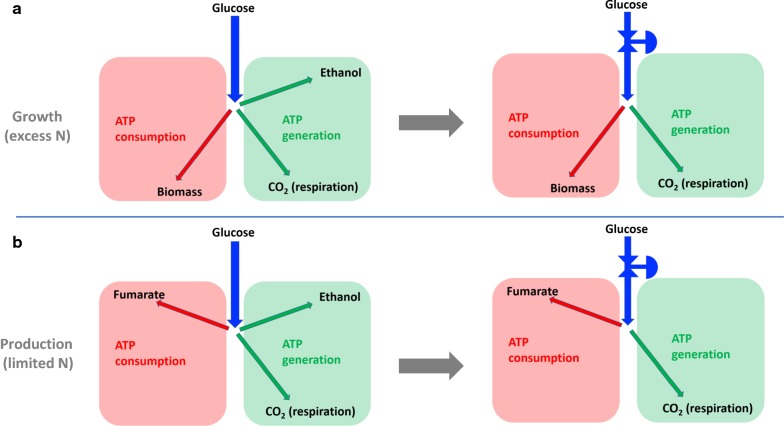
三羧酸循环中的四碳二羧酸(苹果酸、富马酸和琥珀酸)仍然是源自化石原料的各种前体化学品有前景的生物基替代品。富马酸中的双键,除了两个末端羧基外,还开启了一系列下游反应可能性,其中石化衍生的马来酸酐的替代选择值得一提。迄今为止,生产富马酸最有前景的生物体是(ATCC 20344,也称为),它在氮限制条件下自然分泌富马酸。中富马酸的分泌总是与乙醇的共同分泌相关,乙醇是发酵产生的不需要的代谢产物。传统上消除乙醇生产的尝试主要集中在提高菌丝体基质内的氧气可用性。在本研究中,我们采用固定化工艺来研究和利用该生物体的克氏特性,以确定生长和非生长条件下乙醇副产物形成的限度。
所有发酵均在氮过量(生长阶段)或氮限制(生产阶段)下进行,在生长阶段和生产阶段之间进行培养基更换。初始实验在生长和生产阶段均使用过量葡萄糖,同时氧分压在溶解氧为18.4%至85%之间变化。在生长和生产阶段均形成乙醇,氧分压对反应没有影响。结果清楚地表明,菌丝体内可能的厌氧区不是乙醇形成的原因,这表明乙醇是在完全有氧条件下作为代谢溢流产物形成的。对于像这样的克氏阳性生物体,乙醇溢流可通过控制发酵中的葡萄糖输入来控制。在生长和生产发酵中对采用了相同的策略。结果表明,对于葡萄糖添加速率为的情况,在生长过程中所有乙醇都可以消除。生产阶段表现出类似的方式,葡萄糖添加量为导致富马酸产量为,富马酸对葡萄糖的产率为。对葡萄糖添加效果的进一步研究表明,当葡萄糖添加速率达到对生物量时,乙醇溢流开始,而最大葡萄糖摄取速率确定在0.426至之间。
结果确凿地证明是一种克氏阳性生物体,并且该特性可用于完全消除乙醇副产物的形成。展示了一种称为“同型富马酸生产”的状态,其中所有碳输入以富马酸或呼吸的形式离开细胞。基于生物量的最高“同型富马酸生产”:速率为,葡萄糖产率为,这表明了开发无乙醇工艺的界限。本研究中采用的控制策略与固定化工艺简单的可扩展性相结合,为进一步开发生物富马酸生产提供了新方向。