Bioinformatics and Computational Biology Program, University of Minnesota, Minneapolis, MN, USA.
Department of Agronomy and Plant Genetics, University of Minnesota, 1991 Upper Buford Circle, 411 Borlaug Hall, Saint Paul, MN, 55108, USA.
BMC Biotechnol. 2020 Feb 24;20(1):10. doi: 10.1186/s12896-020-00604-3.
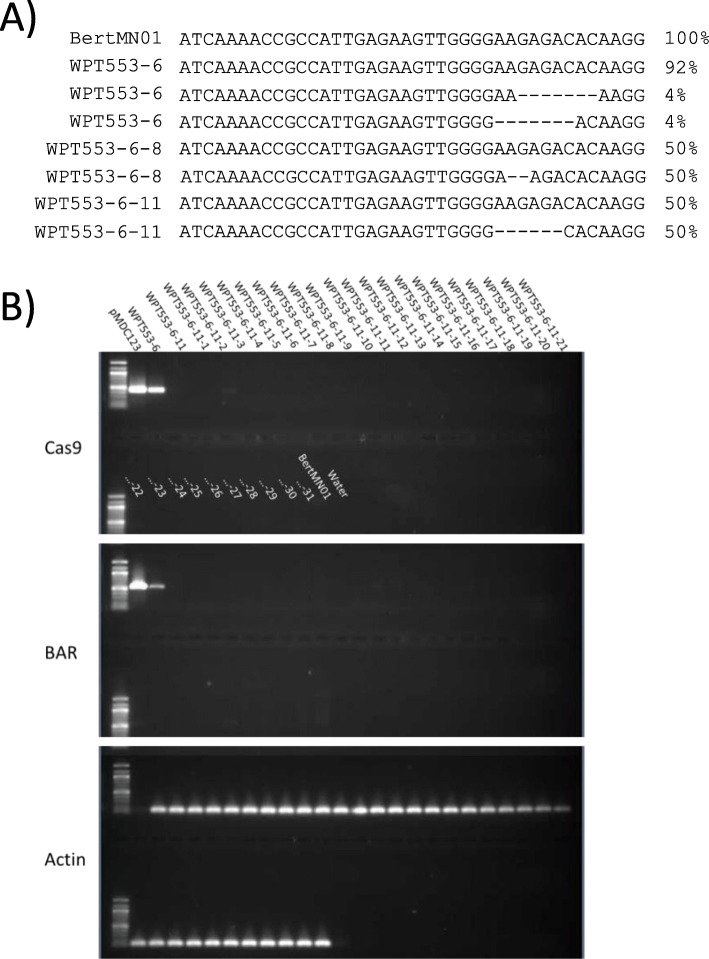
As with many plant species, current genome editing strategies in soybean are initiated by stably transforming a gene that encodes an engineered nuclease into the genome. Expression of the transgene results in a double-stranded break and repair at the targeted locus, oftentimes resulting in mutation(s) at the intended site. As soybean is a self-pollinating species with 20 chromosome pairs, the transgene(s) in the T0 plant are generally expected to be unlinked to the targeted mutation(s), and the transgene(s)/mutation(s) should independently assort into the T1 generation, resulting in Mendellian combinations of transgene presence/absence and allelic states within the segregating family. This prediction, however, is not always consistent with observed results.
In this study, we investigated inheritance patterns among three different CRISPR/Cas9 transgenes and their respective induced mutations in segregating soybean families. Next-generation resequencing of four T0 plants and four T1 progeny plants, followed by broader assessments of the segregating families, revealed both expected and unexpected patterns of inheritance among the different lineages. These unexpected patterns included: (1) A family in which T0 transgenes and mutations were not transmitted to progeny; (2) A family with four unlinked transgene insertions, including two respectively located at paralogous CRISPR target break sites; (3) A family in which mutations were observed and transmitted, but without evidence of transgene integration nor transmission.
Genome resequencing provides high-resolution of transgene integration structures and gene editing events. Segregation patterns of these events can be complicated by several potential mechanisms. This includes, but is not limited to, plant chimeras, multiple unlinked transgene integrations, editing of intended and paralogous targets, linkage between the transgene integration and target site, and transient expression of the editing reagents without transgene integration into the host genome.
与许多植物物种一样,目前大豆的基因组编辑策略是通过稳定转化一个编码工程化核酸酶的基因到基因组中开始的。转基因的表达导致靶向位点的双链断裂和修复,通常导致预期位点的突变(s)。由于大豆是一种自花授粉的 20 对染色体的物种,T0 植物中的转基因(s)通常预计与靶向突变(s)不连锁,并且转基因(s)/突变(s)应该独立地分配到 T1 代中,导致转基因存在/缺失和等位基因状态的孟德尔组合在分离的家系中。然而,这一预测并不总是与观察到的结果一致。
在这项研究中,我们研究了三个不同的 CRISPR/Cas9 转基因及其在分离的大豆家系中诱导的突变的遗传模式。对四个 T0 植物和四个 T1 后代植物进行下一代重测序,然后对分离的家系进行更广泛的评估,揭示了不同谱系之间既有预期的也有意外的遗传模式。这些意外的模式包括:(1)一个家系中 T0 转基因和突变没有传递给后代;(2)一个家系中有四个不连锁的转基因插入,包括分别位于同源 CRISPR 靶位断裂点的两个;(3)一个家系中观察到并传递了突变,但没有转基因整合或传递的证据。
基因组重测序提供了转基因整合结构和基因编辑事件的高分辨率。这些事件的分离模式可能会受到几种潜在机制的影响。这包括但不限于植物嵌合体、多个不连锁的转基因整合、预期和同源靶标的编辑、转基因整合与靶位点之间的连锁以及编辑试剂的瞬时表达而不将其整合到宿主基因组中。