Friedrich Miescher Institute for Biomedical Research, Basel, Switzerland.
University of Basel, Basel, Switzerland.
Elife. 2020 Mar 30;9:e53387. doi: 10.7554/eLife.53387.
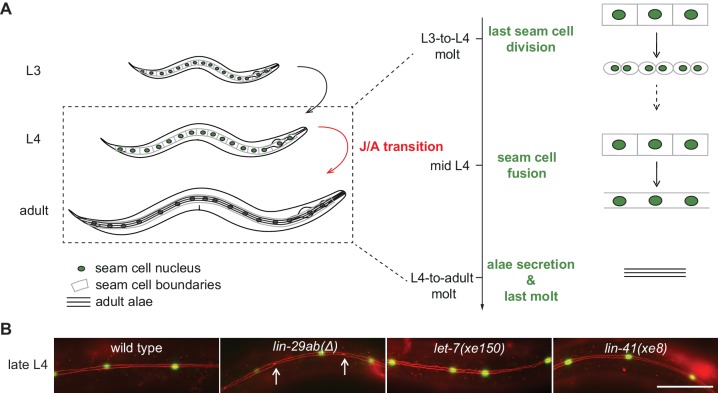
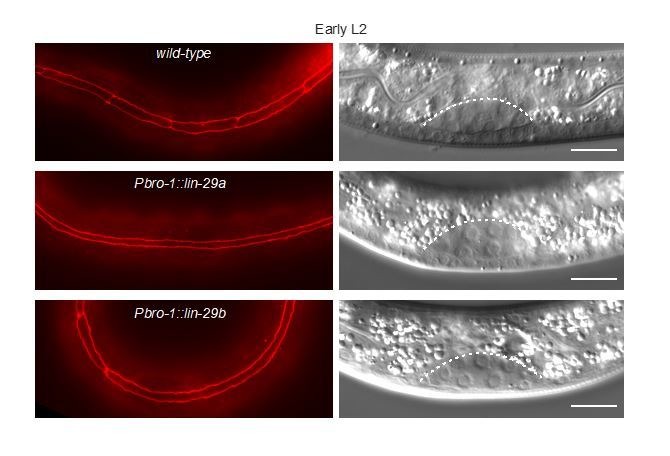
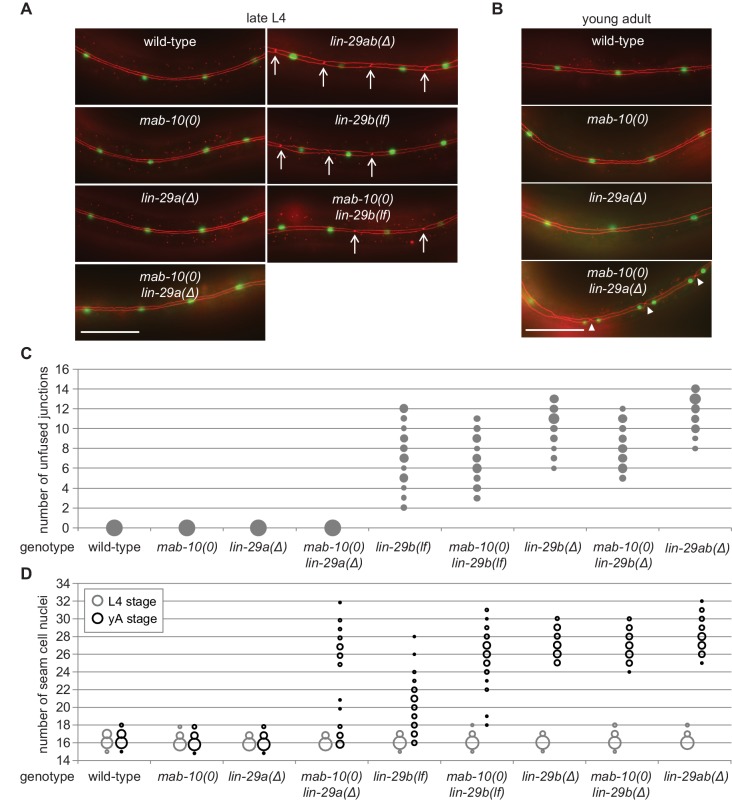
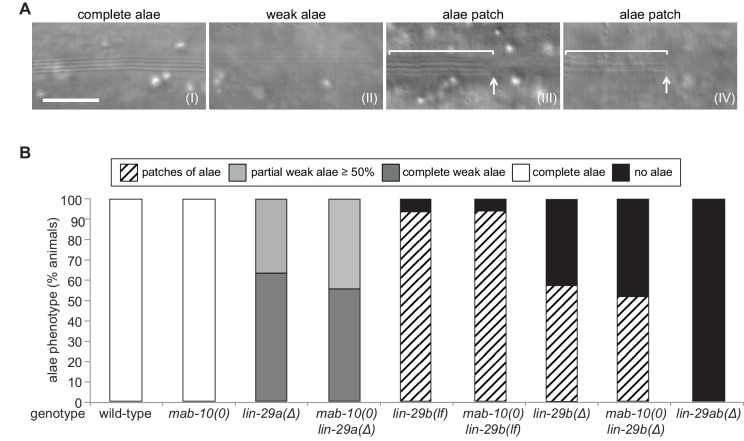
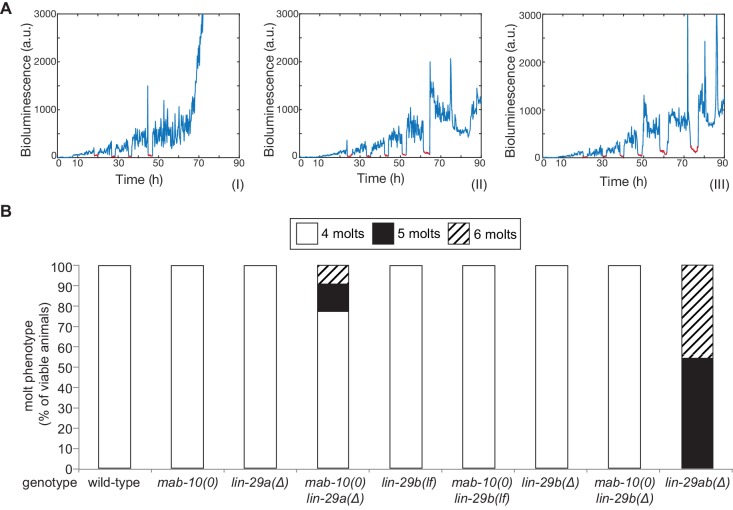
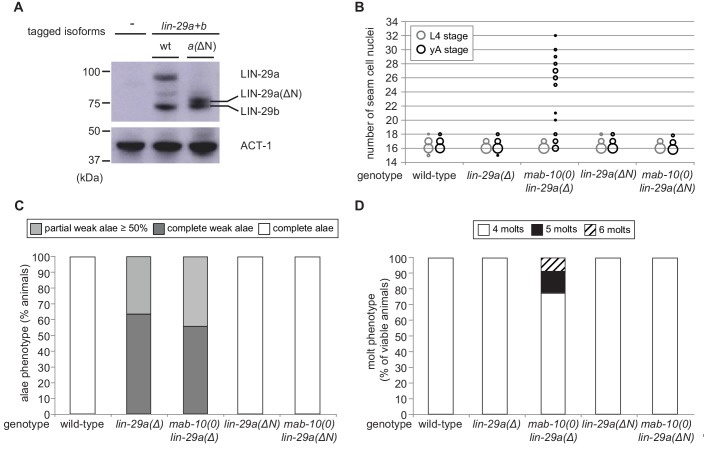
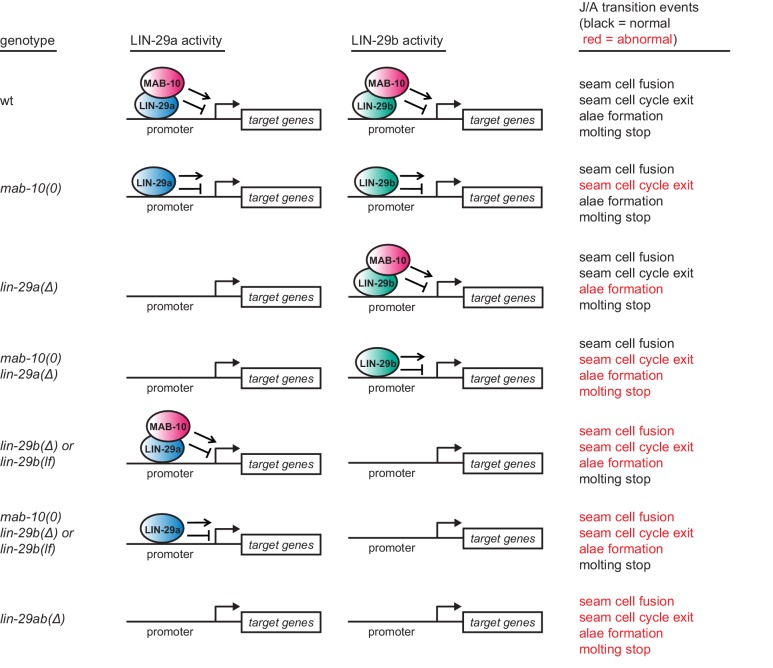
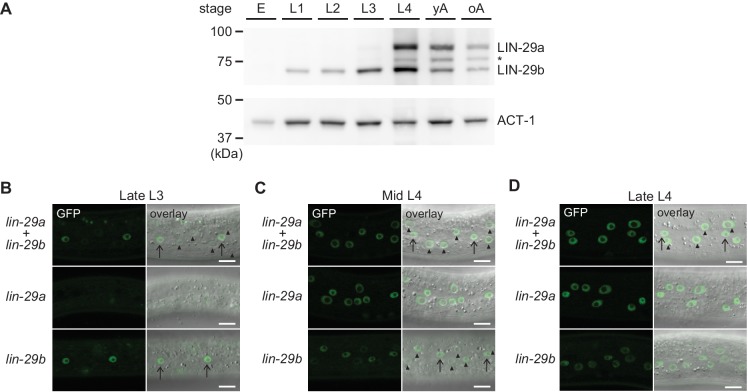
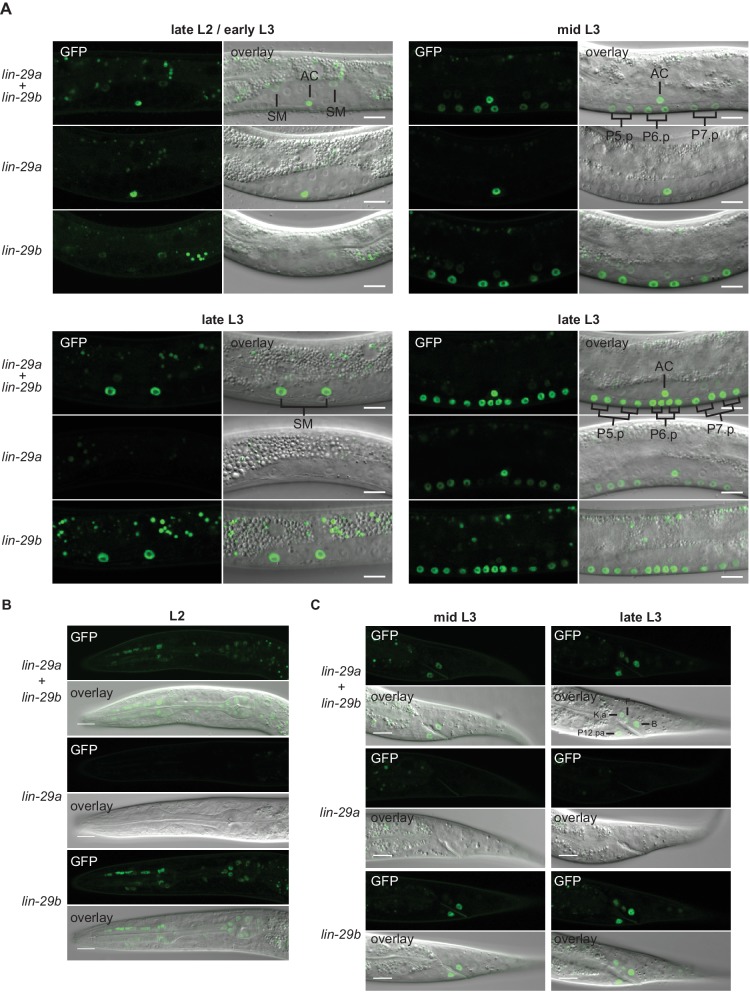
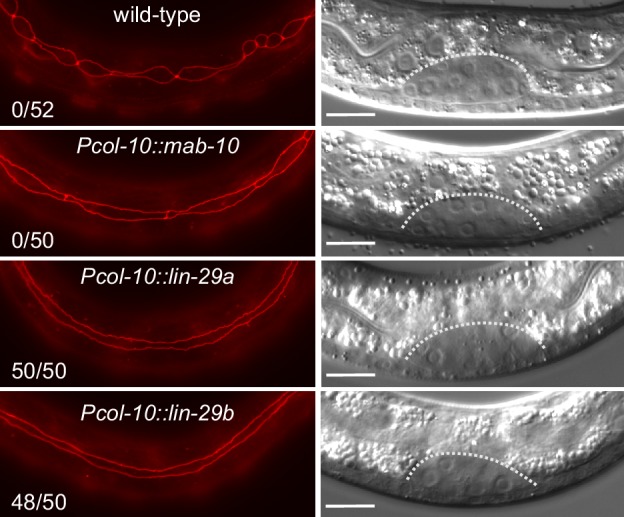
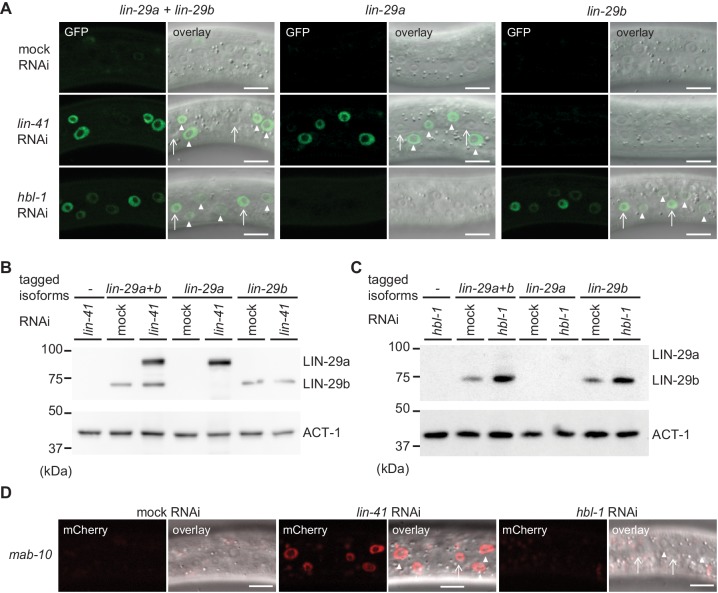
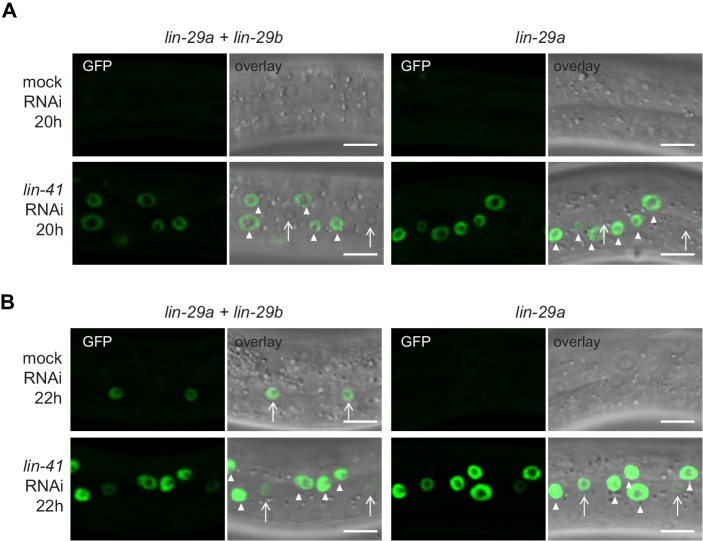
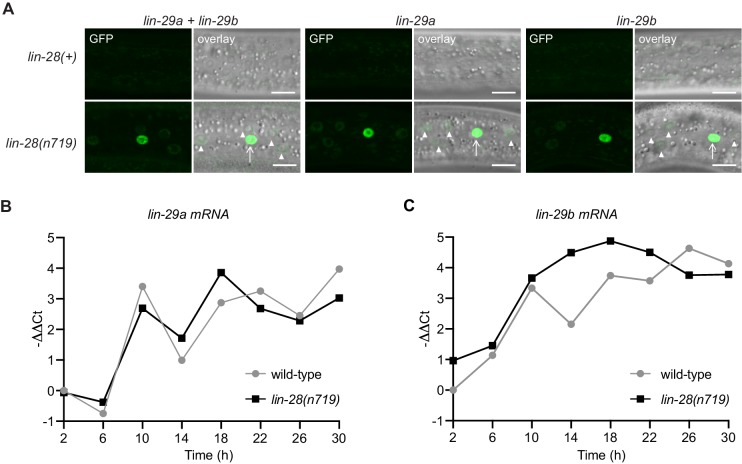
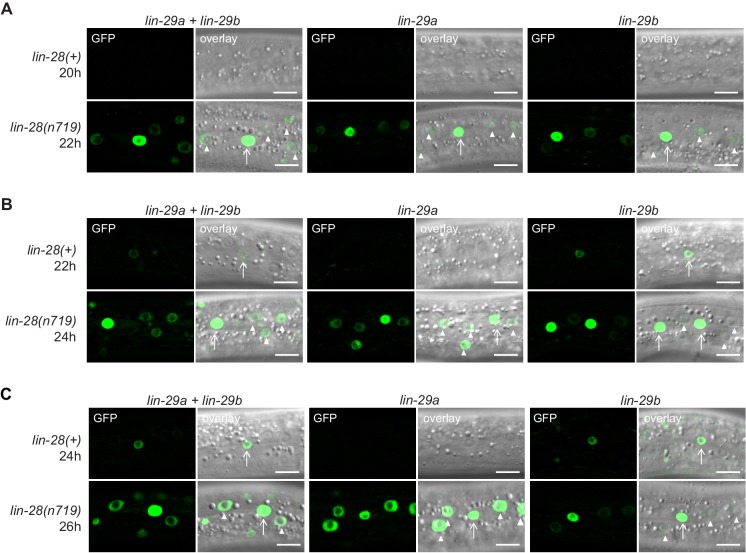
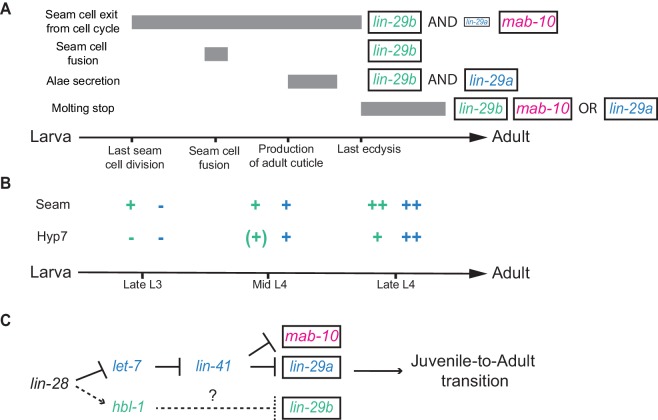
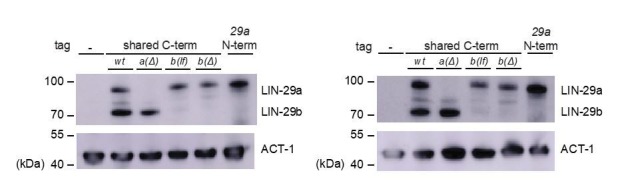
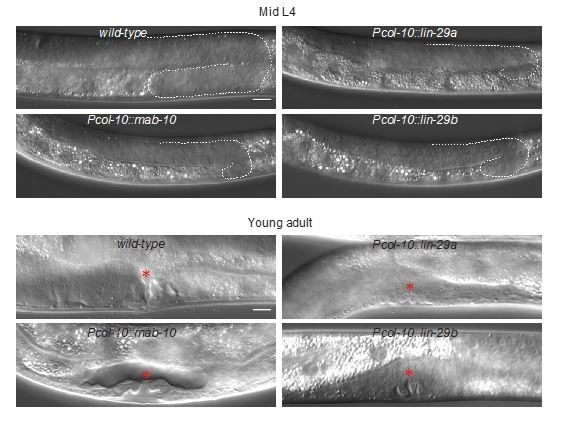
Robust organismal development relies on temporal coordination of disparate physiological processes. In , the heterochronic pathway controls a timely juvenile-to-adult (J/A) transition. This regulatory cascade of conserved proteins and small RNAs culminates in accumulation of the transcription factor LIN-29, which triggers coordinated execution of transition events. We report that two LIN-29 isoforms fulfill distinct functions. Functional specialization is a consequence of distinct isoform expression patterns, not protein sequence, and we propose that distinct LIN-29 dose sensitivities of the individual J/A transition events help to ensure their temporal ordering. We demonstrate that unique isoform expression patterns are generated by the activities of LIN-41 for , and of HBL-1 for 29, whereas the RNA-binding protein LIN-28 coordinates LIN-29 isoform activity, in part by regulating both and . Our findings reveal that coordinated transition from juvenile to adult involves branching of a linear pathway to achieve timely control of multiple events.
生物体的发育需要依赖不同生理过程的时间协调。在 中,异时性途径控制着从幼年到成年(J/A)的适时转变。这一保守蛋白和小 RNA 的调控级联反应最终导致转录因子 LIN-29 的积累,从而引发转变事件的协调执行。我们报告称,两种 LIN-29 同工型具有不同的功能。功能特化是不同同工型表达模式的结果,而不是蛋白质序列的结果,我们提出,个体 J/A 转变事件的不同 LIN-29 剂量敏感性有助于确保它们的时间顺序。我们证明,独特的同工型表达模式是由 LIN-41 为 生成的,由 HBL-1 为 29 生成的,而 RNA 结合蛋白 LIN-28 则通过调节 和 来协调 LIN-29 同工型的活性。我们的发现表明,从幼年到成年的协调转变涉及到线性途径的分支,以实现对多个事件的及时控制。