Friedrich Miescher Institute for Biomedical Research, Basel, Switzerland.
Department of Biosystems Science and Engineering, ETH Zurich, Basel, Switzerland.
Nat Cell Biol. 2021 Nov;23(11):1163-1175. doi: 10.1038/s41556-021-00776-w. Epub 2021 Nov 4.
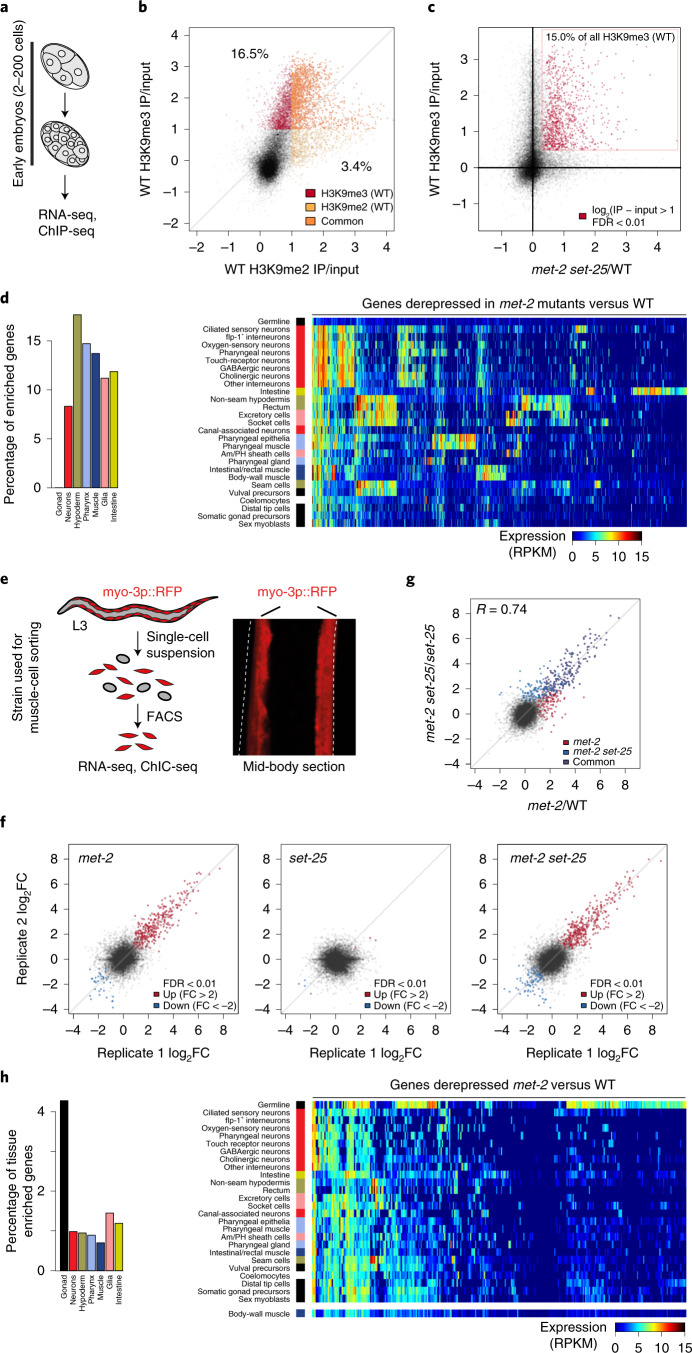
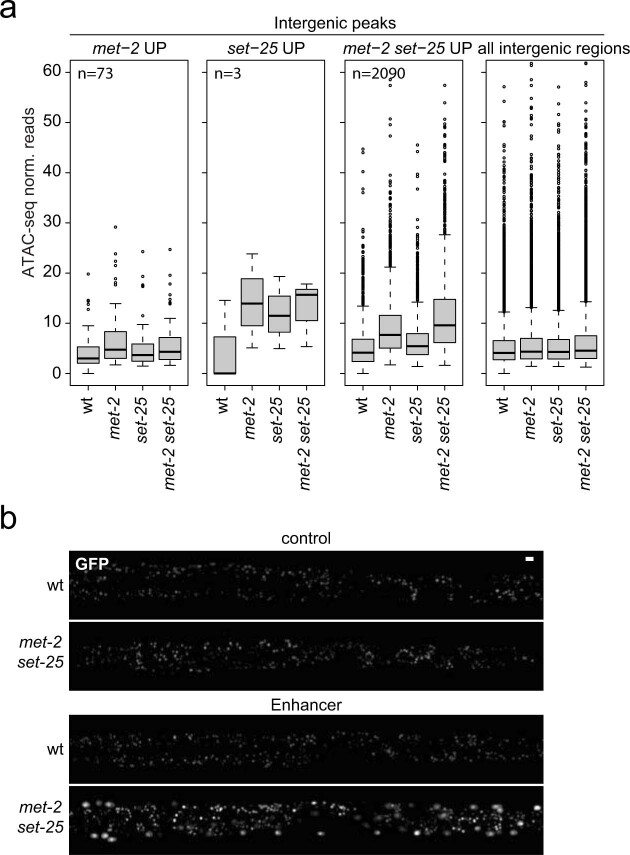
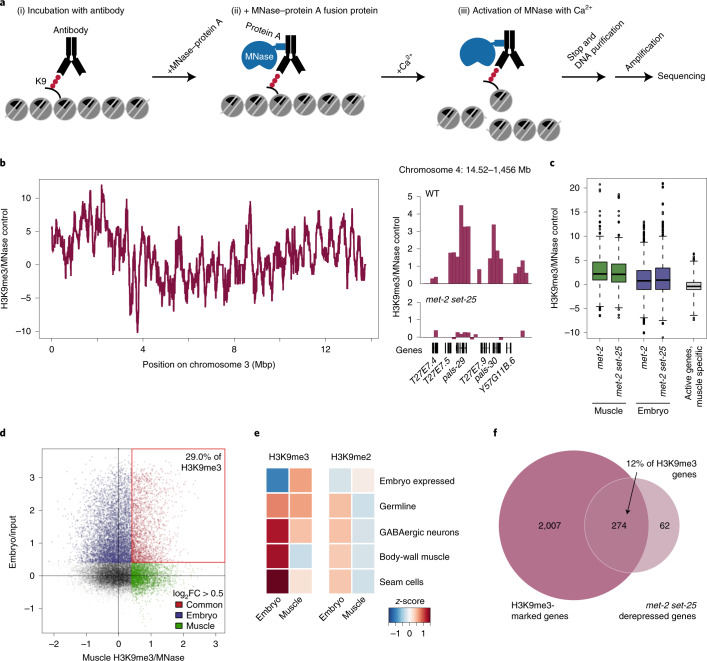
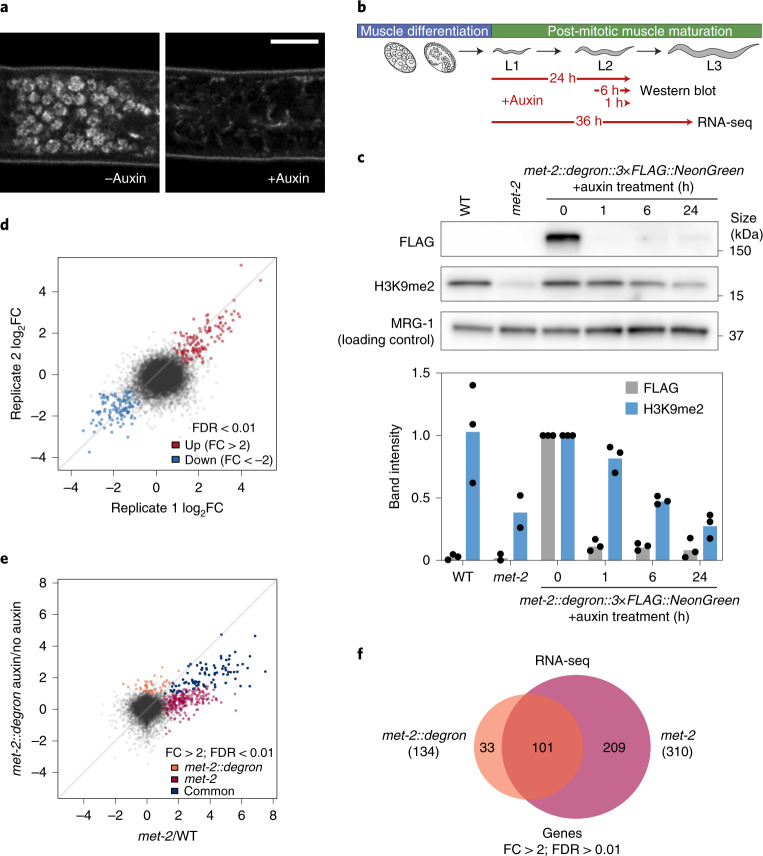
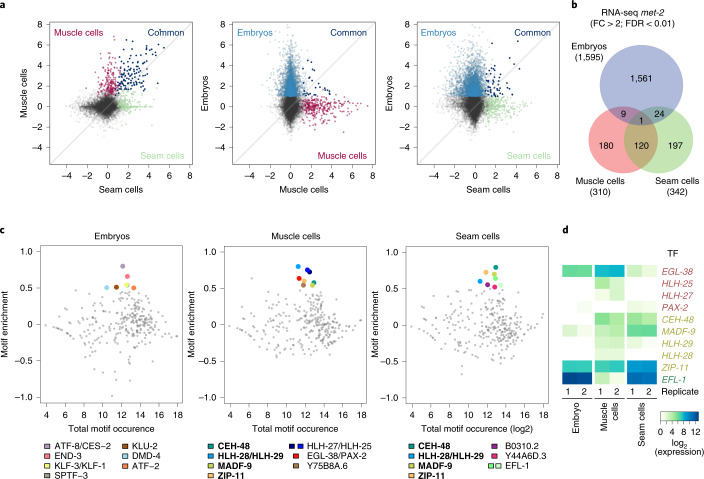
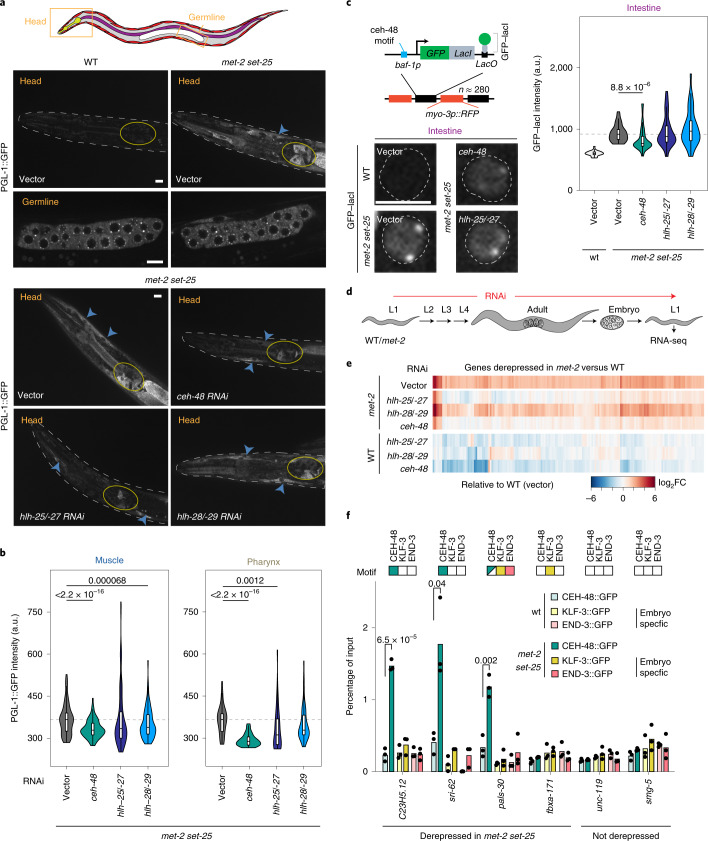
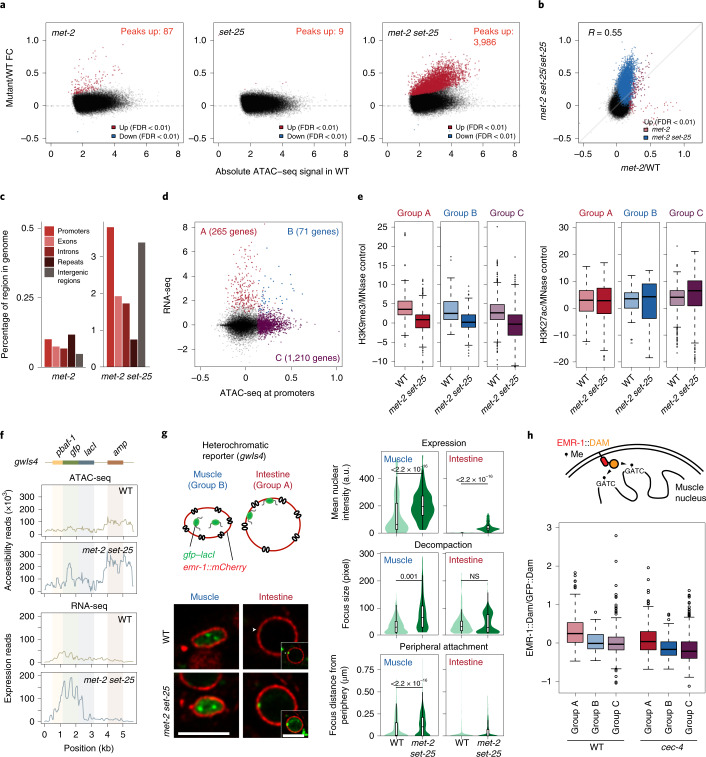
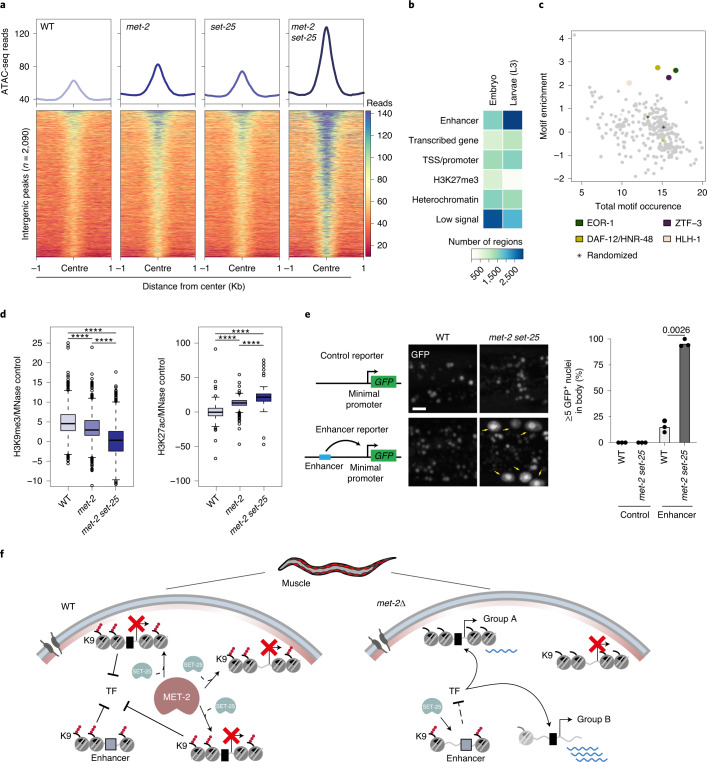
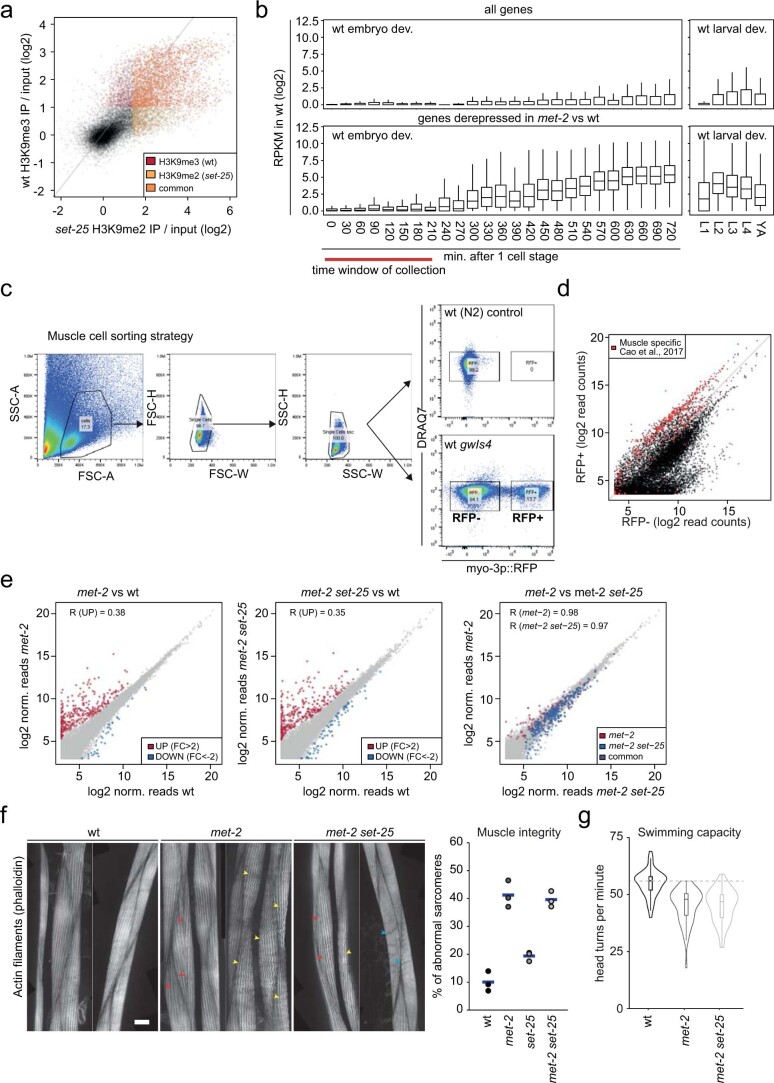
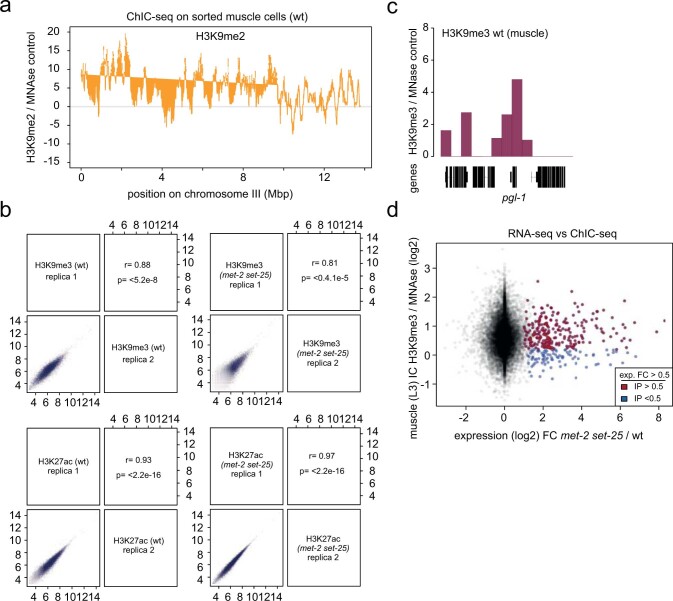
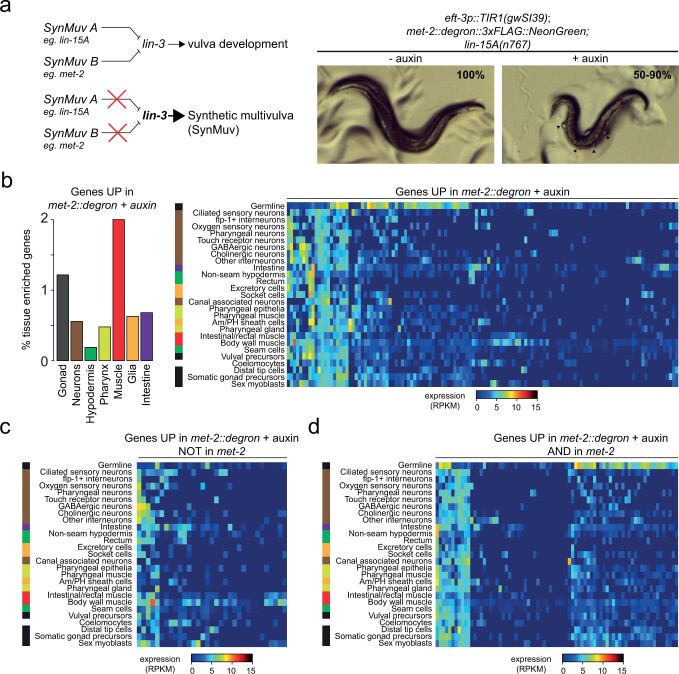
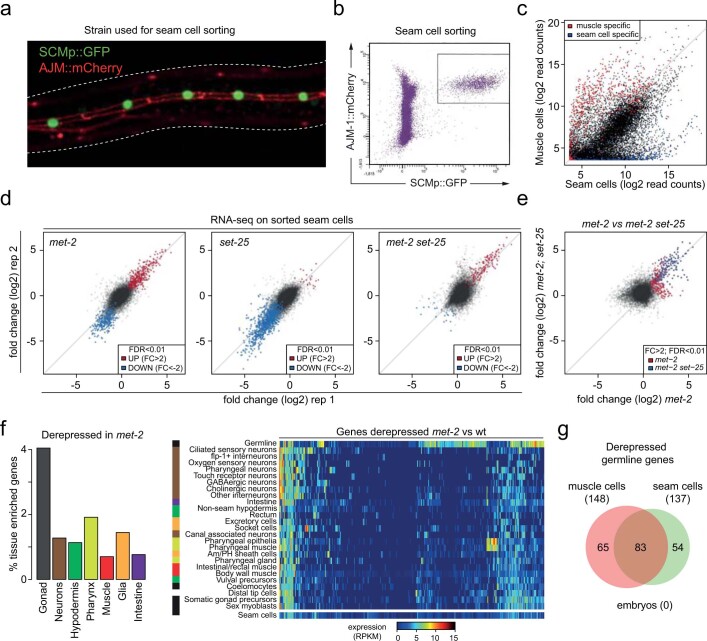
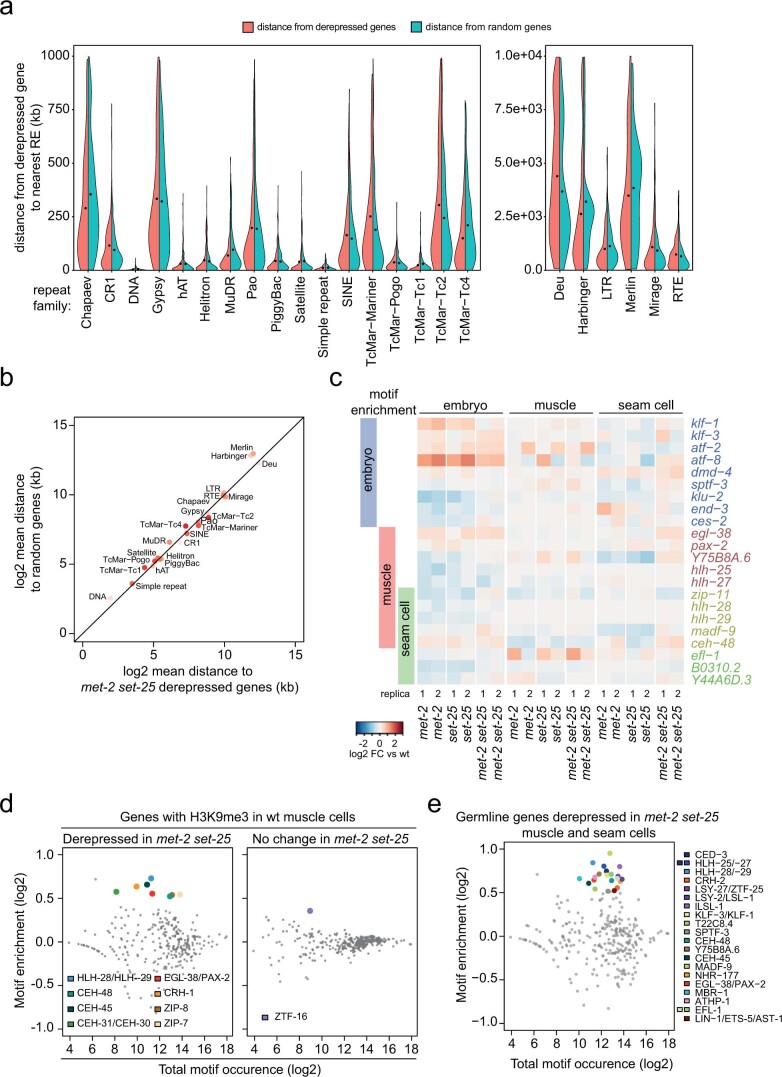
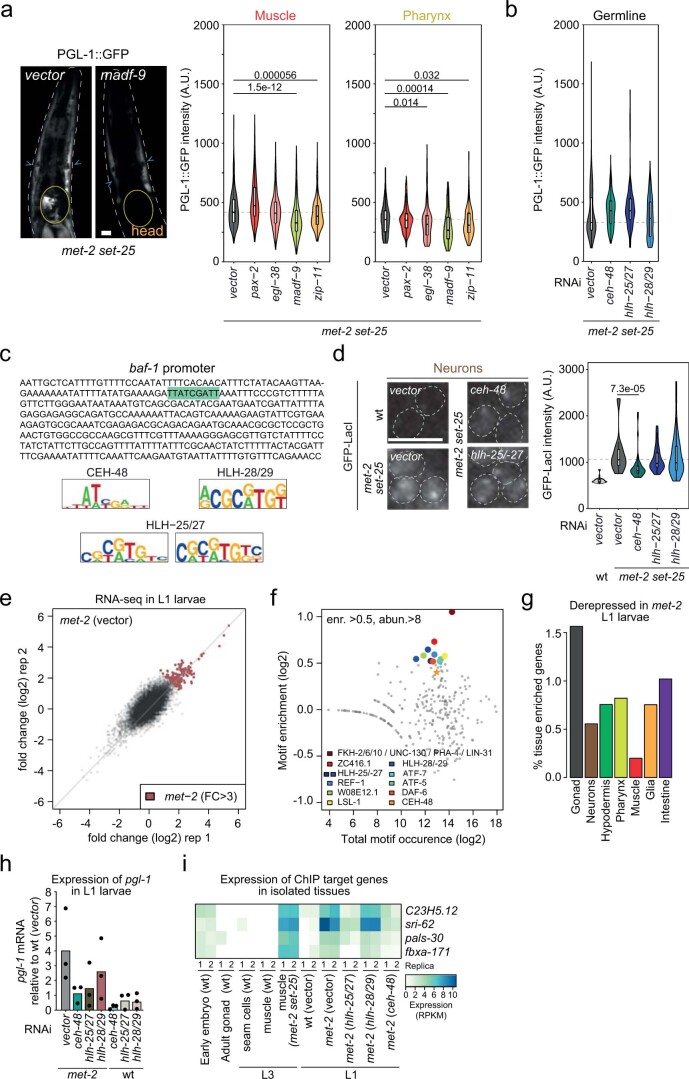
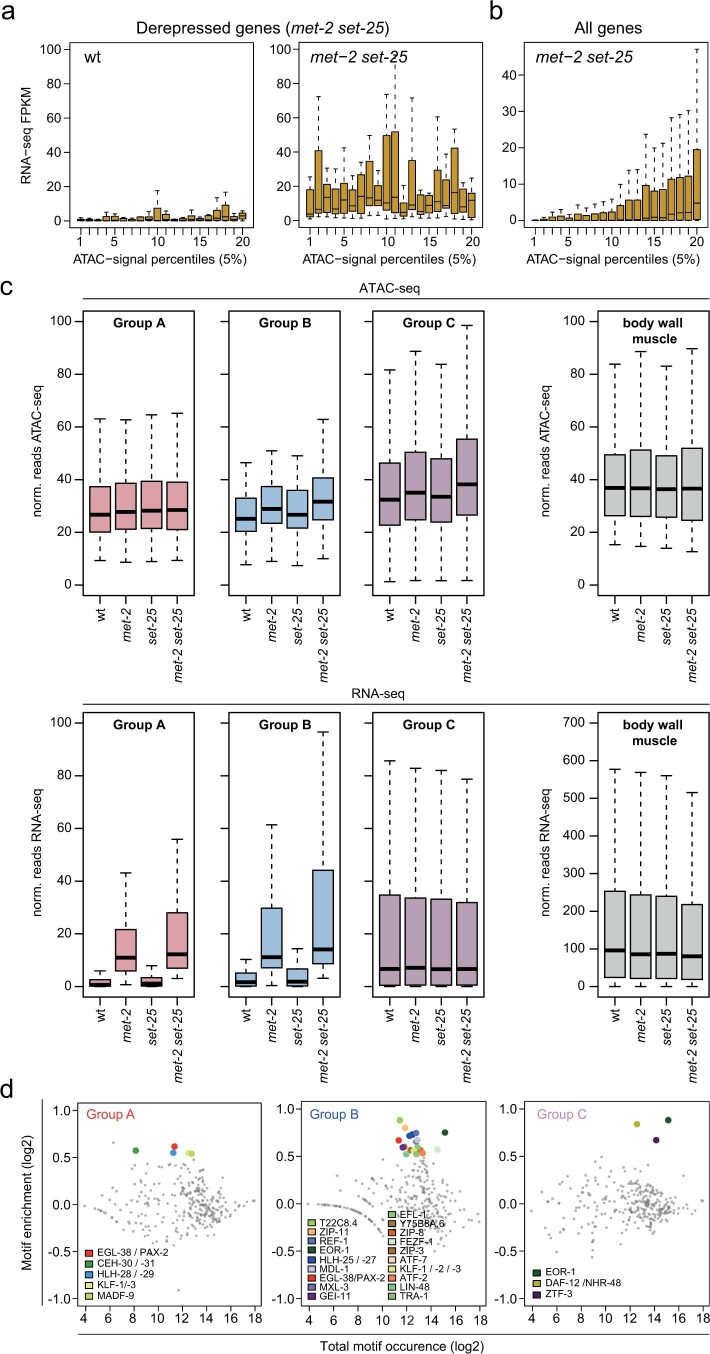
The developmental role of histone H3K9 methylation (H3K9me), which typifies heterochromatin, remains unclear. In Caenorhabditis elegans, loss of H3K9me leads to a highly divergent upregulation of genes with tissue and developmental-stage specificity. During development H3K9me is lost from differentiated cell type-specific genes and gained at genes expressed in earlier developmental stages or other tissues. The continuous deposition of H3K9me2 by the SETDB1 homolog MET-2 after terminal differentiation is necessary to maintain repression. In differentiated tissues, H3K9me ensures silencing by restricting the activity of a defined set of transcription factors at promoters and enhancers. Increased chromatin accessibility following the loss of H3K9me is neither sufficient nor necessary to drive transcription. Increased ATAC-seq signal and gene expression correlate at a subset of loci positioned away from the nuclear envelope, while derepressed genes at the nuclear periphery remain poorly accessible despite being transcribed. In conclusion, H3K9me deposition can confer tissue-specific gene expression and maintain the integrity of terminally differentiated muscle by restricting transcription factor activity.
组蛋白 H3K9 甲基化(H3K9me)的发育作用尚不清楚,其特征是异染色质。在秀丽隐杆线虫中,H3K9me 的缺失导致具有组织和发育阶段特异性的基因高度差异上调。在发育过程中,H3K9me 从分化细胞类型特异性基因中丢失,并在早期发育阶段或其他组织中表达的基因中获得。SETDB1 同源物 MET-2 在后分化过程中持续沉积 H3K9me2 对于维持抑制是必要的。在分化的组织中,H3K9me 通过限制启动子和增强子处的一组特定转录因子的活性来确保沉默。H3K9me 丢失后染色质可及性的增加既不足以也不需要驱动转录。在远离核膜定位的一部分基因座上,增加的 ATAC-seq 信号和基因表达相关,而核周去抑制的基因尽管转录但仍难以接近。总之,H3K9me 的沉积可以赋予组织特异性基因表达,并通过限制转录因子的活性来维持终末分化肌肉的完整性。