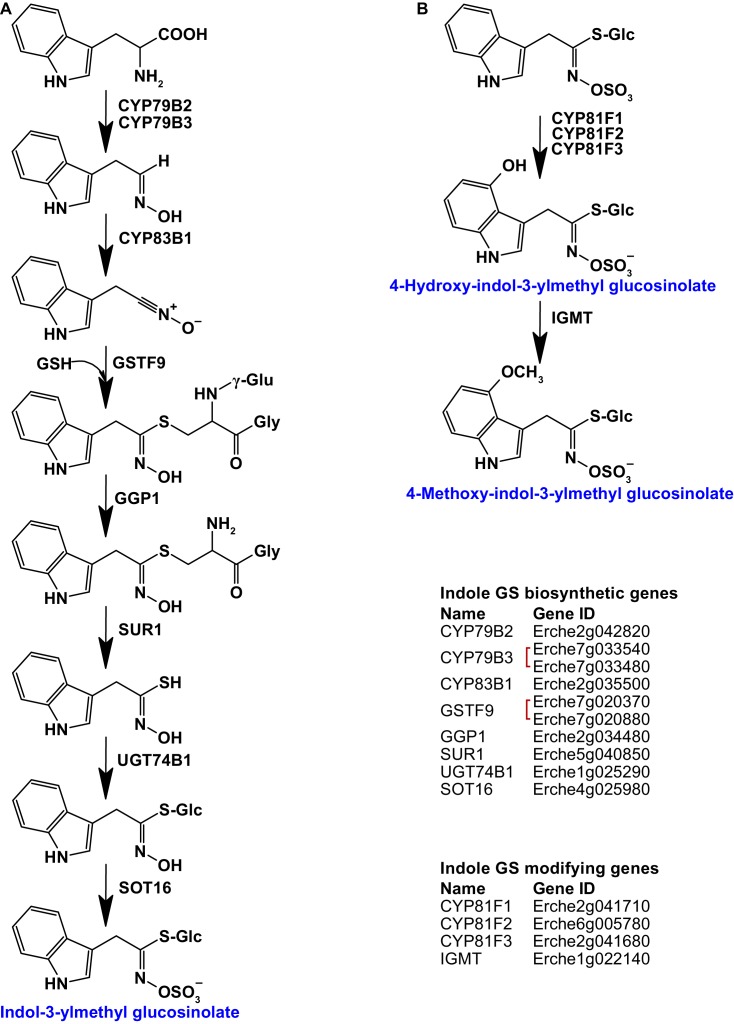
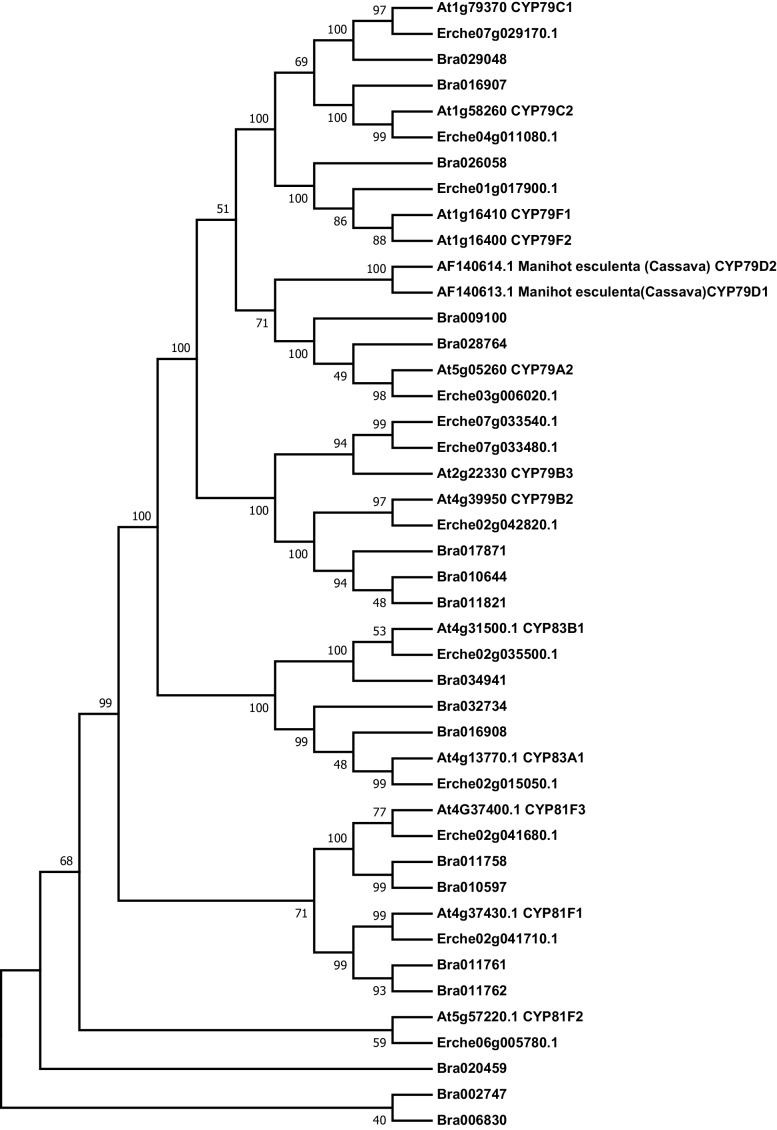
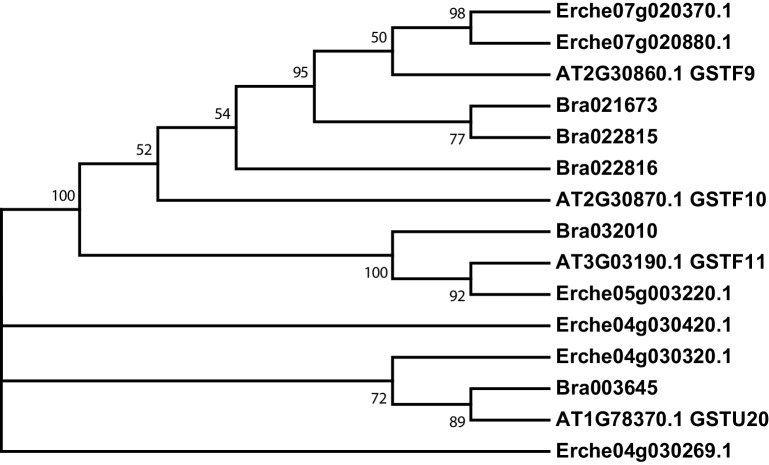
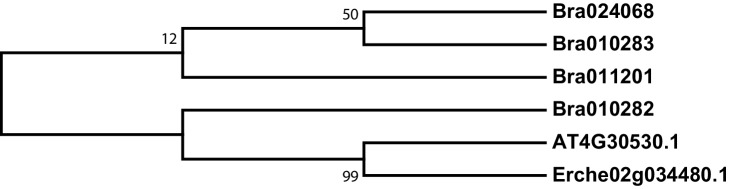
Institute of Plant Sciences, University of Bern, Bern, Switzerland.
Boyce Thompson Institute, Ithaca, United States.
Elife. 2020 Apr 7;9:e51712. doi: 10.7554/eLife.51712.
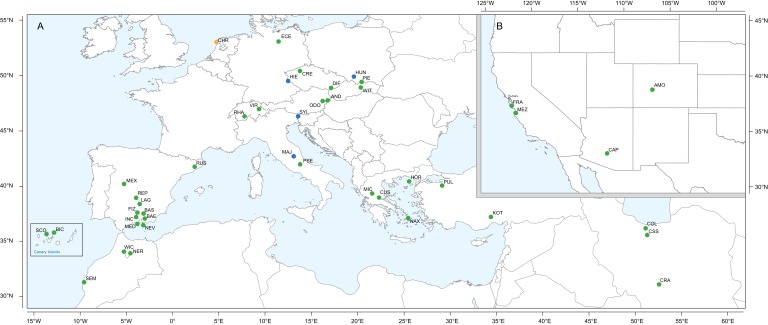
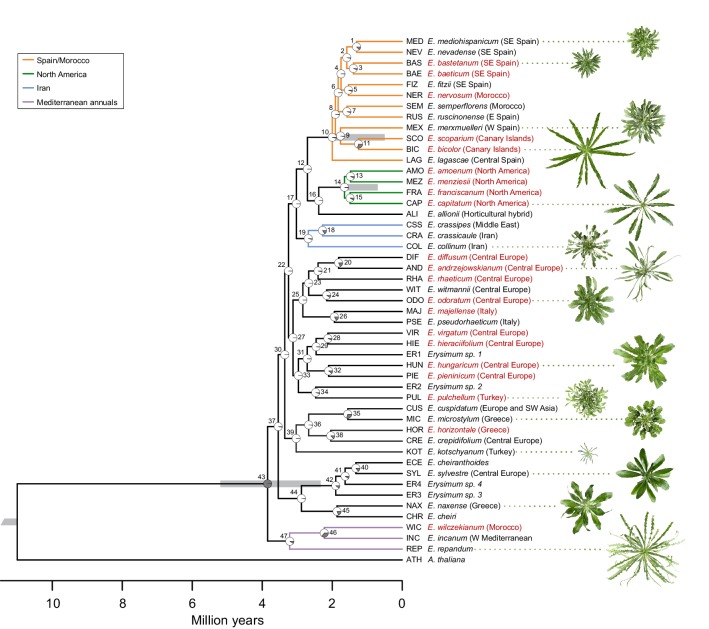
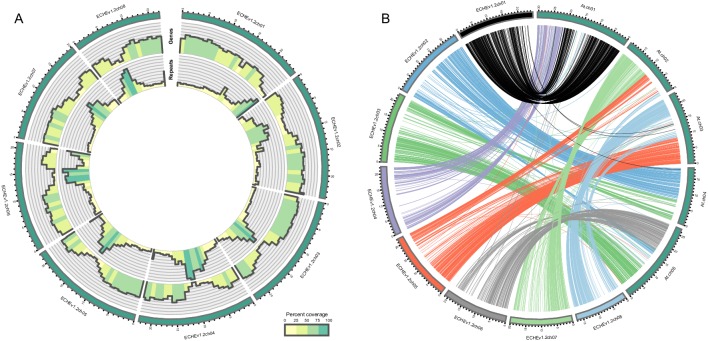
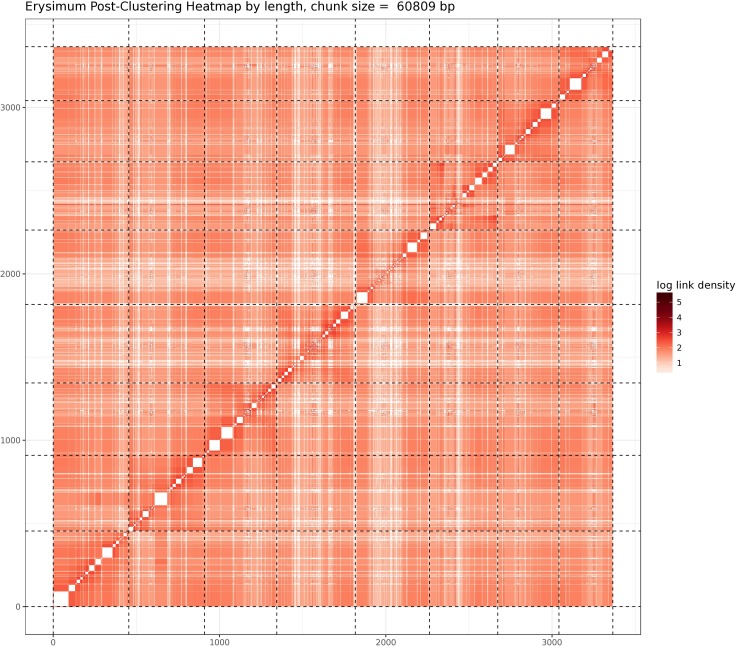
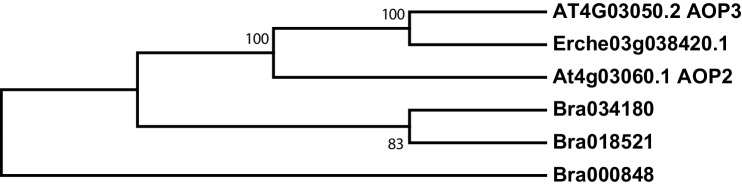
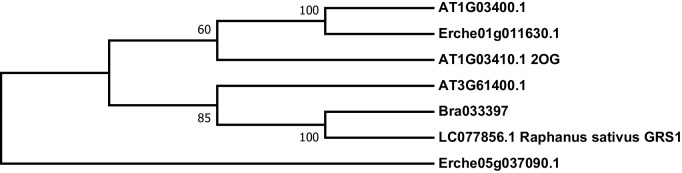
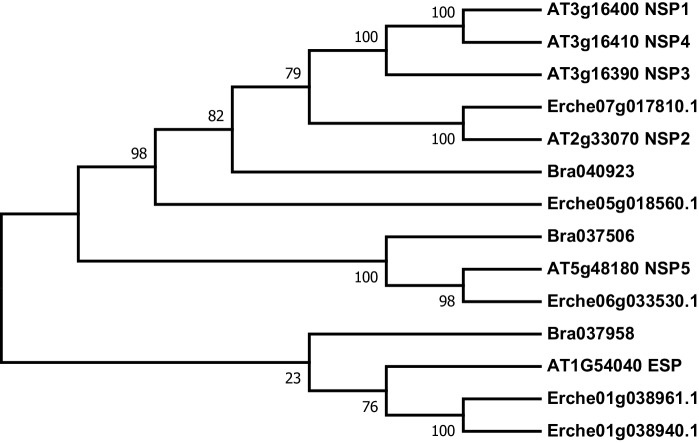
Phytochemical diversity is thought to result from coevolutionary cycles as specialization in herbivores imposes diversifying selection on plant chemical defenses. Plants in the speciose genus (Brassicaceae) produce both ancestral glucosinolates and evolutionarily novel cardenolides as defenses. Here we test macroevolutionary hypotheses on co-expression, co-regulation, and diversification of these potentially redundant defenses across this genus. We sequenced and assembled the genome of and foliar transcriptomes of 47 additional species to construct a phylogeny from 9868 orthologous genes, revealing several geographic clades but also high levels of gene discordance. Concentrations, inducibility, and diversity of the two defenses varied independently among species, with no evidence for trade-offs. Closely related, geographically co-occurring species shared similar cardenolide traits, but not glucosinolate traits, likely as a result of specific selective pressures acting on each defense. Ancestral and novel chemical defenses in thus appear to provide complementary rather than redundant functions.
植物化学多样性被认为是协同进化循环的结果,因为草食动物的特化对植物的化学防御施加了多样化选择。在物种丰富的 属(十字花科)中,植物既产生原始的硫代葡萄糖苷,也产生进化上新颖的卡烯内酯作为防御。在这里,我们测试了这些潜在冗余防御在协同表达、协同调控和多样化方面的宏观进化假设。我们对 进行了测序和组装,并对 47 个附加 物种的叶片转录组进行了测序,构建了一个来自 9868 个直系同源基因的系统发育树,揭示了几个地理分支,但也存在高水平的基因不一致性。两种防御物质的浓度、诱导性和多样性在物种间独立变化,没有证据表明存在权衡。密切相关的、地理上共存的物种具有相似的卡烯内酯特征,但没有硫代葡萄糖苷特征,这可能是由于每种防御物质受到特定的选择性压力。因此, 中的祖先和新的化学防御似乎提供了互补而不是冗余的功能。