The Pirbright Institute, Woking, United Kingdom.
Section of Virology, Faculty of Medicine, Imperial College London, London, United Kingdom.
J Virol. 2020 Jun 16;94(13). doi: 10.1128/JVI.02107-19.
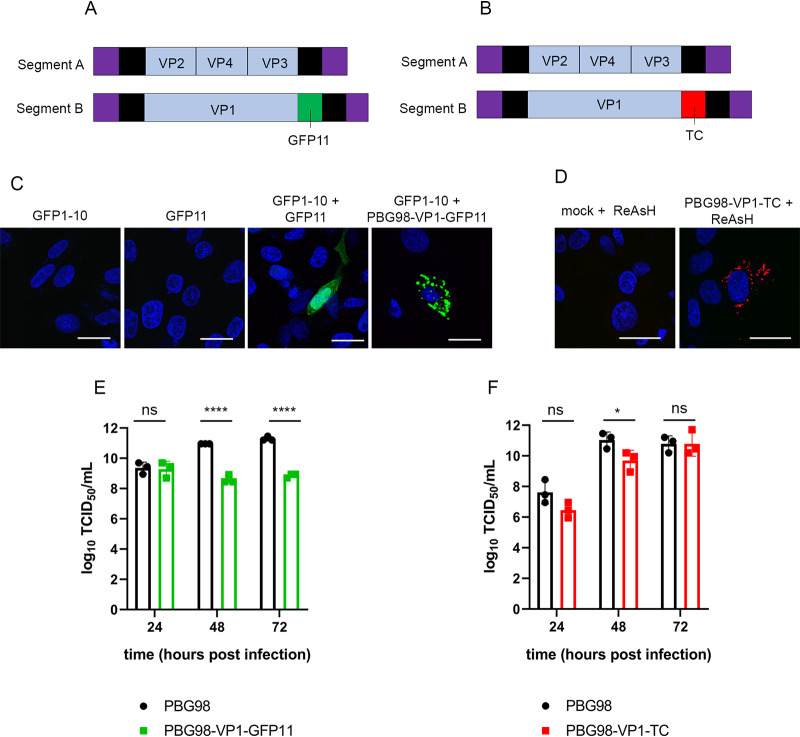
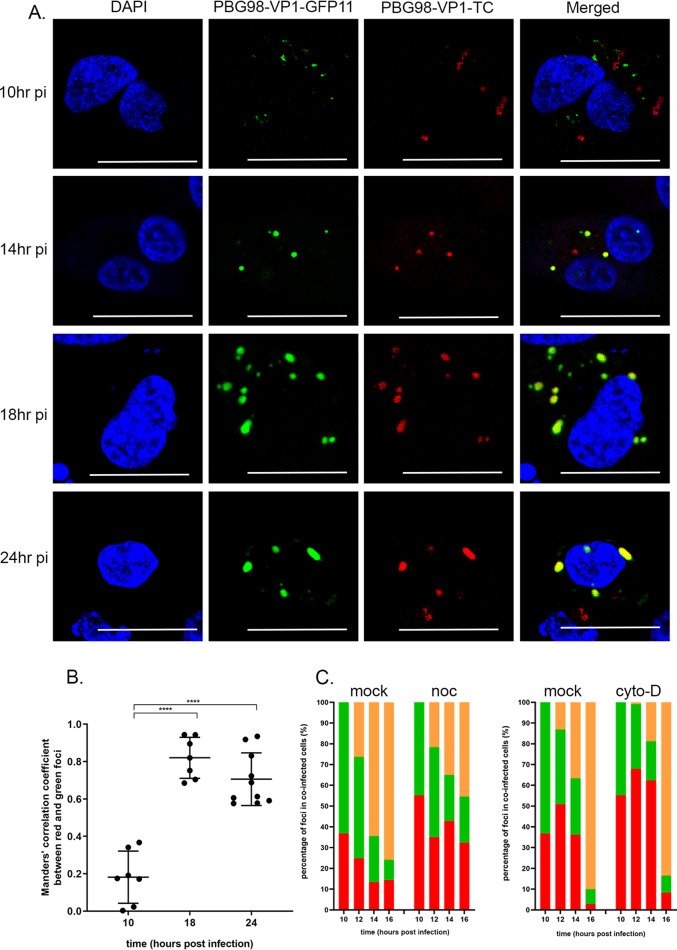
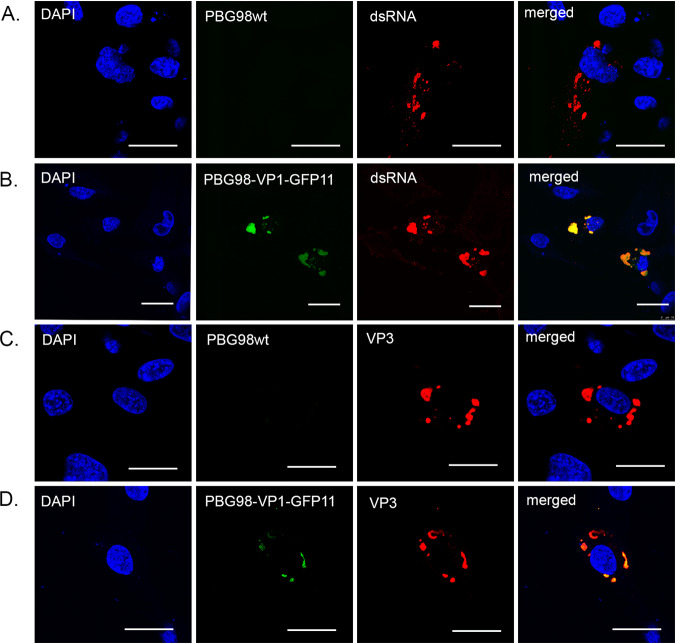
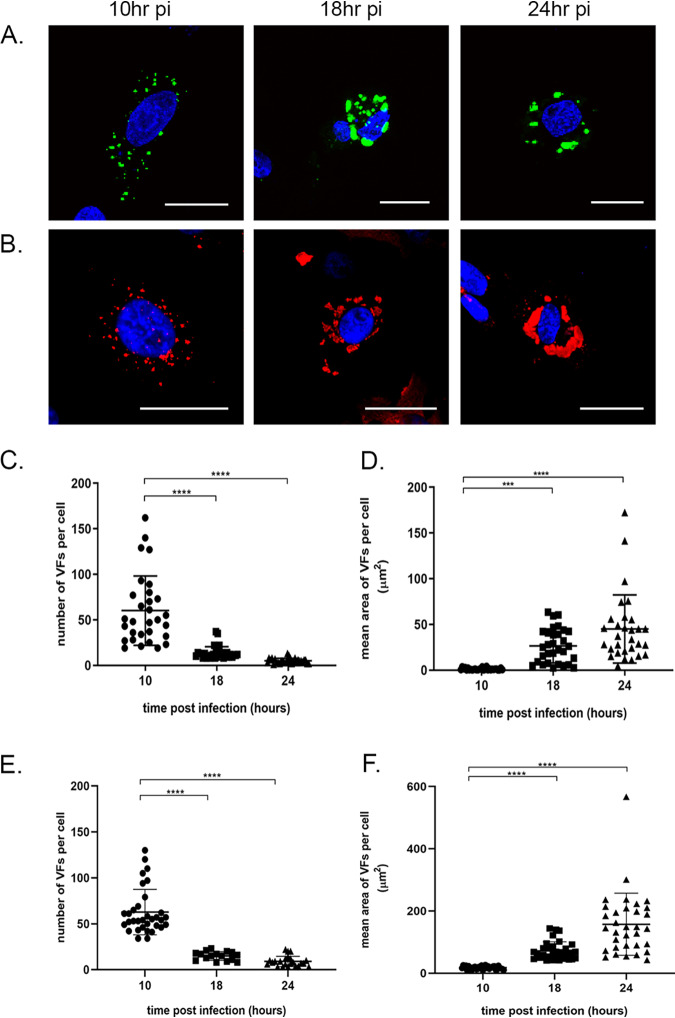
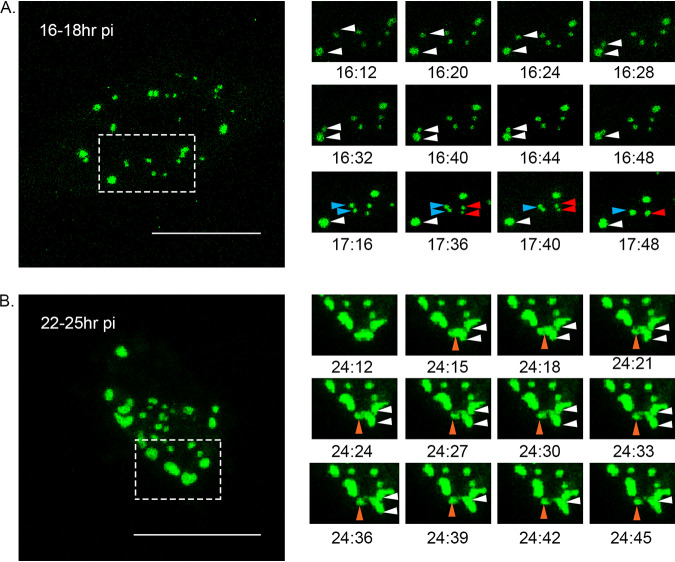
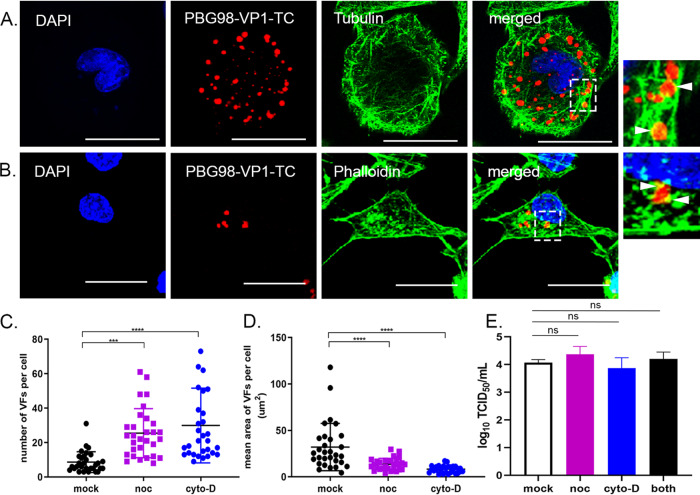
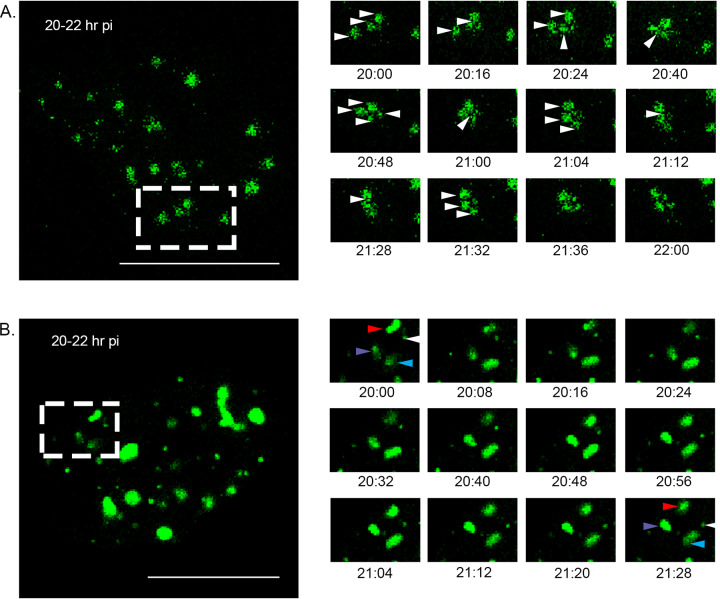
The family, responsible for major economic losses to poultry and aquaculture, is composed of nonenveloped viruses with a segmented double-stranded RNA (dsRNA) genome that replicate in discrete cytoplasmic virus factories (VFs). Reassortment is common; however, the underlying mechanism remains unknown given that VFs may act as a barrier to genome mixing. In order to provide new information on VF trafficking during dsRNA virus coinfection, we rescued two recombinant infectious bursal disease viruses (IBDVs) of strain PBG98 containing either a split GFP11 or a tetracysteine (TC) tag fused to the VP1 polymerase (PBG98-VP1-GFP11 and PBG98-VP1-TC). DF-1 cells transfected with GFP1-10 prior to PBG98-VP1-GFP11 infection or stained with a biarsenical derivative of the red fluorophore resorufin (ReAsH) following PBG98-VP1-TC infection, had green or red foci in the cytoplasm, respectively, that colocalized with VP3 and dsRNA, consistent with VFs. The average number of VFs decreased from a mean of 60 to 5 per cell between 10 and 24 h postinfection (hpi) ( < 0.0001), while the average area increased from 1.24 to 45.01 μm ( < 0.0001), and live cell imaging revealed that the VFs were highly dynamic structures that coalesced in the cytoplasm. Small VFs moved faster than large (average 0.57 μm/s at 16 hpi compared to 0.22 μm/s at 22 hpi), and VF coalescence was dependent on an intact microtubule network and actin cytoskeleton. During coinfection with PBG98-VP1-GFP11 and PBG98-VP1-TC viruses, discrete VFs initially formed from each input virus that subsequently coalesced 10 to 16 hpi, and we speculate that reassortment requires VF coalescence. Reassortment is common in viruses with segmented double-stranded RNA (dsRNA) genomes. However, these viruses typically replicate within discrete cytoplasmic virus factories (VFs) that may represent a barrier to genome mixing. We generated the first replication competent tagged reporter birnaviruses, infectious bursal disease viruses (IBDVs) containing a split GFP11 or tetracysteine (TC) tag and used the viruses to track the location and movement of IBDV VFs, in order to better understand the intracellular dynamics of VFs during a coinfection. Discrete VFs initially formed from each virus that subsequently coalesced from 10 h postinfection. We hypothesize that VF coalescence is required for the reassortment of the This study provides new information that adds to our understanding of dsRNA virus VF trafficking.
家族,对家禽和水产养殖造成重大经济损失,由具有分段双链 RNA(dsRNA)基因组的无包膜病毒组成,该基因组在离散的细胞质病毒工厂(VF)中复制。重配很常见;然而,由于 VF 可能是基因组混合的障碍,因此其潜在机制尚不清楚。为了提供 dsRNA 病毒共感染期间 VF 运输的新信息,我们拯救了两种重组传染性法氏囊病病毒(IBDV)株 PBG98,其中包含融合到 VP1 聚合酶(PBG98-VP1-GFP11 和 PBG98-VP1-TC)的分裂 GFP11 或四半胱氨酸(TC)标签。在用 PBG98-VP1-GFP11 感染之前用 GFP1-10 转染的 DF-1 细胞中,或者在用 PBG98-VP1-TC 感染后用红色荧光团 resorufin 的生物偶联衍生物(ReAsH)染色,细胞质中分别具有绿色或红色焦点,与 VP3 和 dsRNA 共定位,与 VF 一致。在感染后 10 至 24 小时(hpi)之间,VF 的平均数量从每细胞 60 个减少到 5 个(<0.0001),而平均面积从 1.24 增加到 45.01 μm(<0.0001),活细胞成像显示 VF 是高度动态的结构,在细胞质中融合。小 VF 比大 VF 移动得更快(在 16 hpi 时平均为 0.57 μm/s,而在 22 hpi 时为 0.22 μm/s),VF 融合依赖于完整的微管网络和肌动蛋白细胞骨架。在 PBG98-VP1-GFP11 和 PBG98-VP1-TC 病毒共感染期间,来自每个输入病毒的离散 VF 最初形成,随后在 10 至 16 hpi 时融合,我们推测重配需要 VF 融合。具有分段双链 RNA(dsRNA)基因组的病毒中重配很常见。然而,这些病毒通常在离散的细胞质病毒工厂(VF)内复制,这可能是基因组混合的障碍。我们生成了第一个具有复制能力的标记报告双 RNA 病毒,传染性法氏囊病病毒(IBDV)含有分裂 GFP11 或四半胱氨酸(TC)标签,并使用这些病毒来跟踪 IBDV VF 的位置和运动,以便更好地了解共感染期间 VF 的细胞内动力学。来自每个病毒的离散 VF 最初形成,随后在感染后 10 小时融合。我们假设 VF 融合是重配的必要条件。这项研究提供了新的信息,增加了我们对 dsRNA 病毒 VF 运输的理解。