Structural Biology Program, Van Andel Institute, Grand Rapids, MI, USA.
Howard Hughes Medical Institute, Chevy Chase, MD, USA.
Nat Commun. 2020 Jun 22;11(1):3156. doi: 10.1038/s41467-020-16910-5.
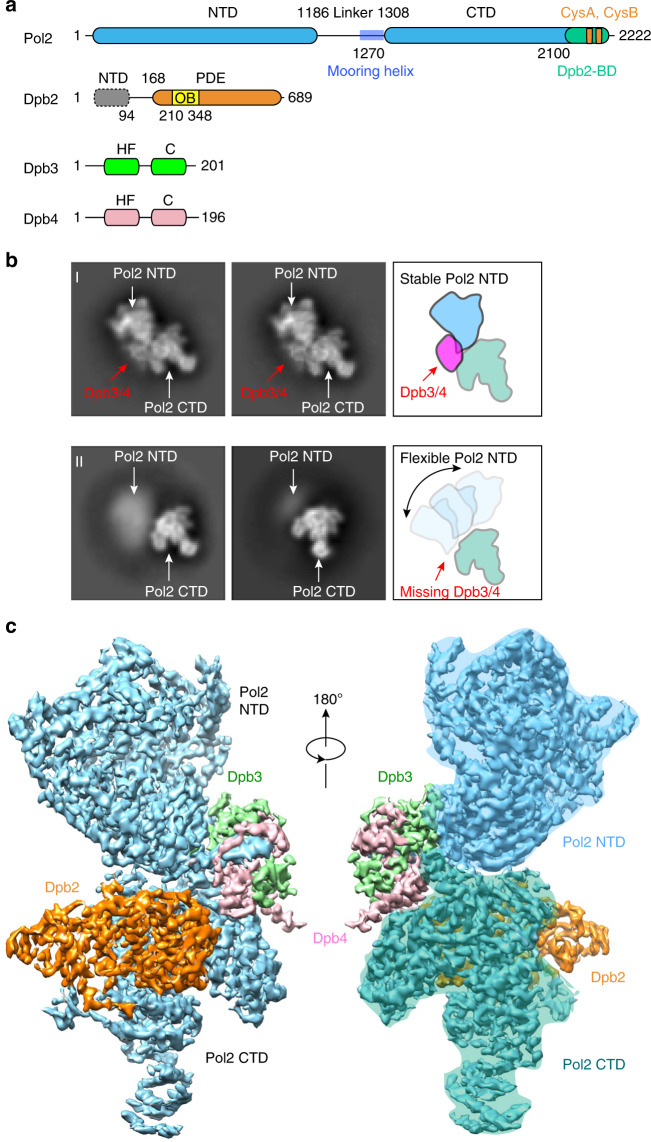
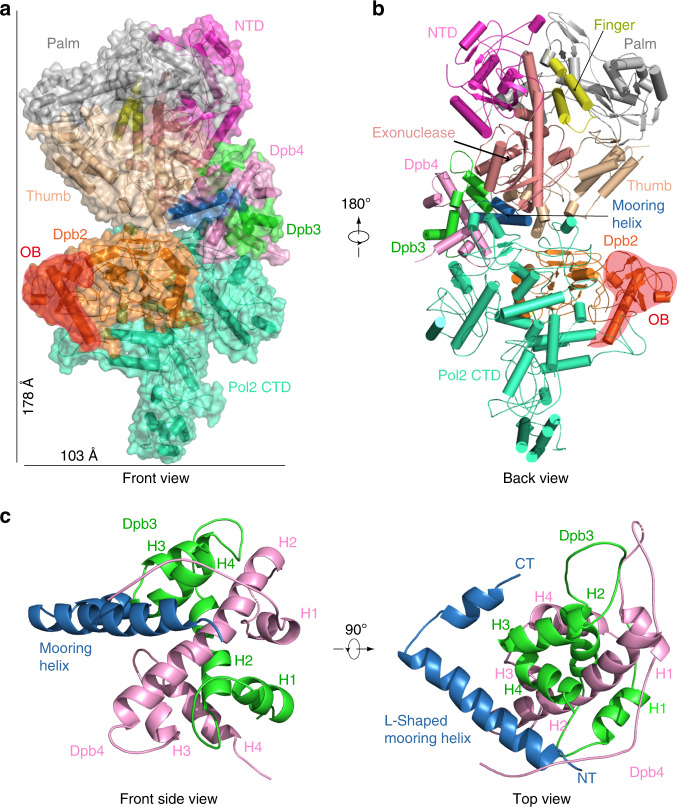
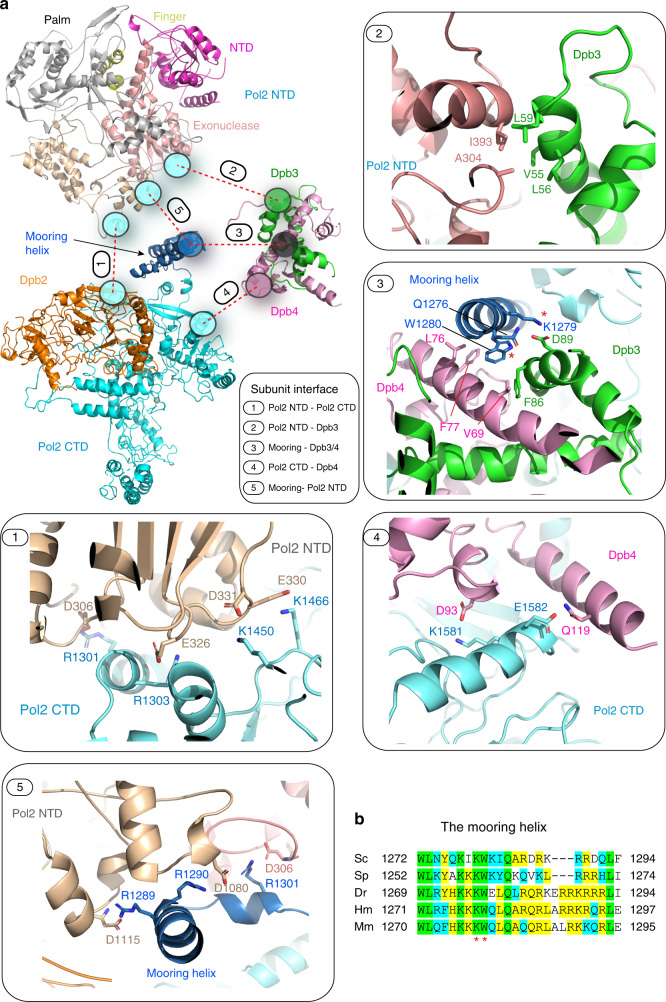
The eukaryotic leading strand DNA polymerase (Pol) ε contains 4 subunits, Pol2, Dpb2, Dpb3 and Dpb4. Pol2 is a fusion of two B-family Pols; the N-terminal Pol module is catalytic and the C-terminal Pol module is non-catalytic. Despite extensive efforts, there is no atomic structure for Pol ε holoenzyme, critical to understanding how DNA synthesis is coordinated with unwinding and the DNA path through the CMG helicase-Pol ε-PCNA clamp. We show here a 3.5-Å cryo-EM structure of yeast Pol ε revealing that the Dpb3-Dpb4 subunits bridge the two DNA Pol modules of Pol2, holding them rigid. This information enabled an atomic model of the leading strand replisome. Interestingly, the model suggests that an OB fold in Dbp2 directs leading ssDNA from CMG to the Pol ε active site. These results complete the DNA path from entry of parental DNA into CMG to exit of daughter DNA from PCNA.
真核生物领头链 DNA 聚合酶 (Pol) ε 包含 4 个亚基,Pol2、Dpb2、Dpb3 和 Dpb4。Pol2 是两个 B 族 Pol 的融合体;N 端 Pol 结构域具有催化活性,而 C 端 Pol 结构域没有催化活性。尽管进行了广泛的努力,但仍没有 Pol ε 全酶的原子结构,这对于理解 DNA 合成如何与解旋以及 DNA 通过 CMG 解旋酶-Pol ε-PCNA 夹的路径相协调至关重要。我们在此展示了酵母 Pol ε 的 3.5 Å 冷冻电镜结构,揭示了 Dpb3-Dpb4 亚基桥接了 Pol2 的两个 DNA Pol 结构域,使它们保持刚性。这些信息使领头链复制体的原子模型成为可能。有趣的是,该模型表明 Dbp2 中的 OB 折叠将领头链 ssDNA 从 CMG 引导至 Pol ε 的活性位点。这些结果完成了从亲本 DNA 进入 CMG 到子 DNA 从 PCNA 退出的 DNA 路径。