Galili Uri
Rush Medical College, Rush University, Chicago, IL 60612, USA.
Int J Mol Sci. 2020 Sep 13;21(18):6702. doi: 10.3390/ijms21186702.
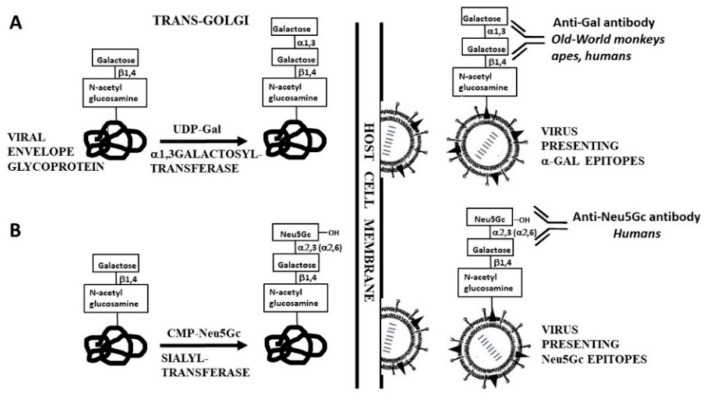
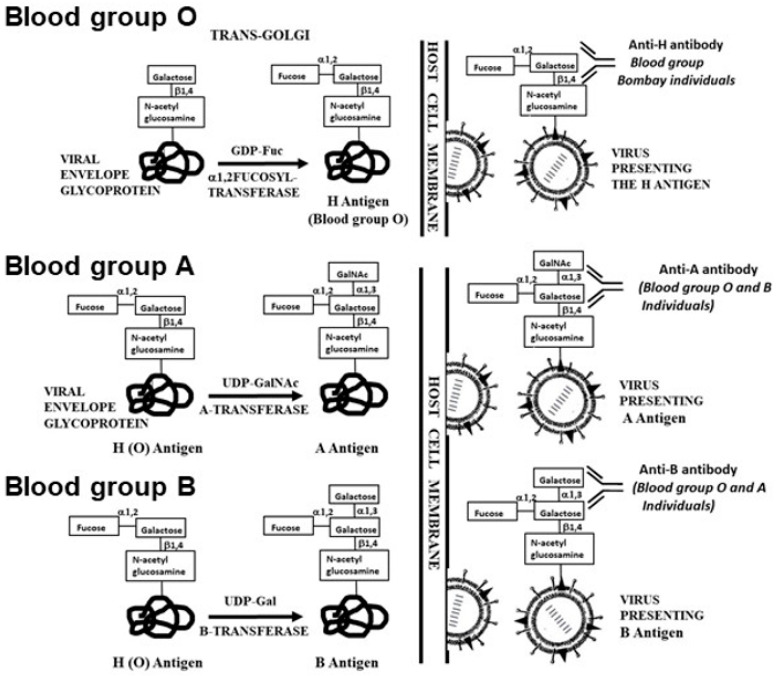
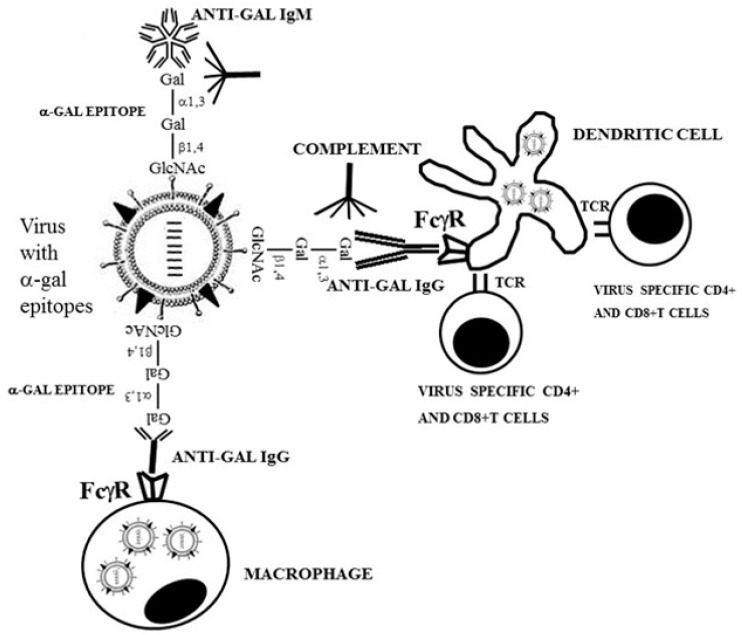
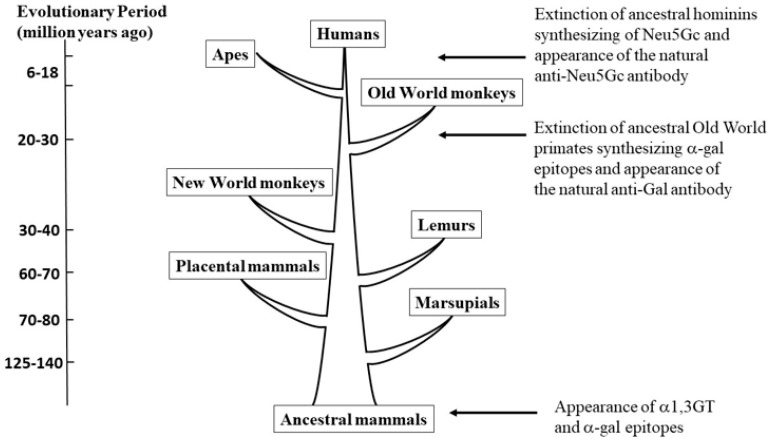
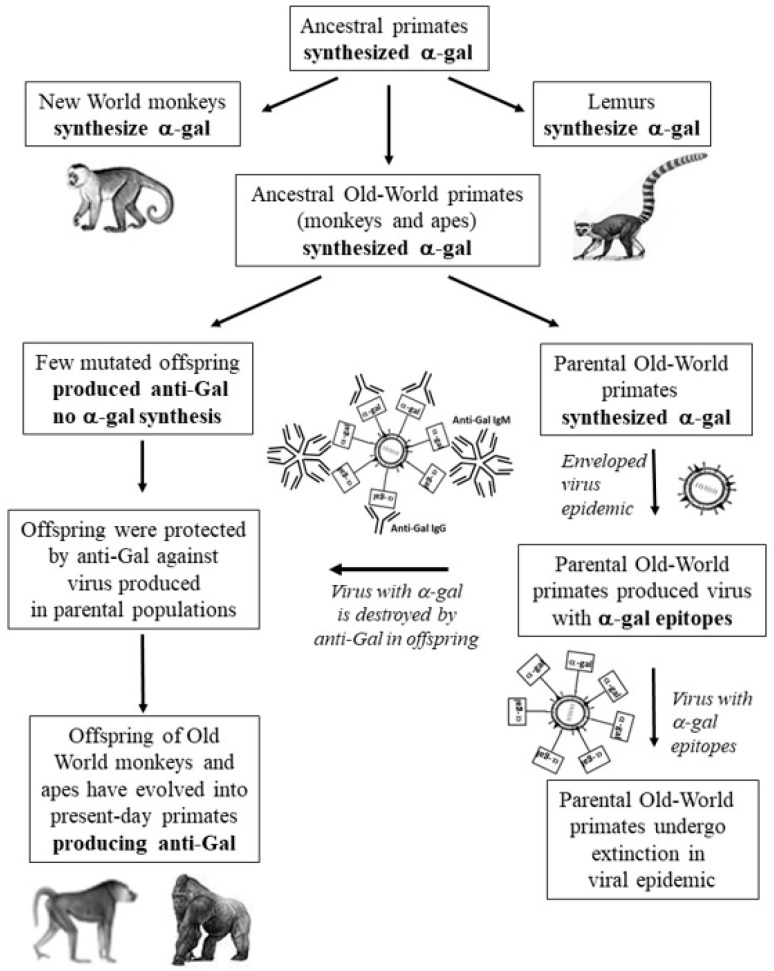
The glycans on enveloped viruses are synthesized by host-cell machinery. Some of these glycans on zoonotic viruses of mammalian reservoirs are recognized by human natural antibodies that may protect against such viruses. These antibodies are produced mostly against carbohydrate antigens on gastrointestinal bacteria and fortuitously, they bind to carbohydrate antigens synthesized in other mammals, neutralize and destroy viruses presenting these antigens. Two such antibodies are: anti-Gal binding to -gal epitopes synthesized in non-primate mammals, lemurs, and New World monkeys, and anti-N-glycolyl neuraminic acid (anti-Neu5Gc) binding to N-glycolyl-neuraminic acid (Neu5Gc) synthesized in apes, Old World monkeys, and many non-primate mammals. Anti-Gal appeared in Old World primates following accidental inactivation of the 1,3galactosyltransferase gene 20-30 million years ago. Anti-Neu5Gc appeared in hominins following the inactivation of the cytidine-monophosphate-N-acetyl-neuraminic acid hydroxylase gene, which led to the loss of Neu5Gc <6 million-years-ago. It is suggested that an epidemic of a lethal virus eliminated ancestral Old World-primates synthesizing -gal epitopes, whereas few mutated offspring lacking -gal epitopes and producing anti-Gal survived because anti-Gal destroyed viruses presenting -gal epitopes, following replication in parental populations. Similarly, anti-Neu5Gc protected few mutated hominins lacking Neu5Gc in lethal virus epidemics that eliminated parental hominins synthesizing Neu5Gc. Since -gal epitopes are presented on many zoonotic viruses it is suggested that vaccines elevating anti-Gal titers may be of protective significance in areas endemic for such zoonotic viruses. This protection would be during the non-primate mammal to human virus transmission, but not in subsequent human to human transmission where the virus presents human glycans. In addition, production of viral vaccines presenting multiple -gal epitopes increases their immunogenicity because of effective anti-Gal-mediated targeting of vaccines to antigen presenting cells for extensive uptake of the vaccine by these cells.
包膜病毒上的聚糖由宿主细胞机制合成。哺乳动物宿主的人畜共患病病毒上的一些聚糖可被人类天然抗体识别,这些抗体可能对这类病毒具有保护作用。这些抗体大多是针对胃肠道细菌上的碳水化合物抗原产生的,偶然情况下,它们会与其他哺乳动物合成的碳水化合物抗原结合,中和并摧毁呈现这些抗原的病毒。两种这样的抗体是:抗Gal,它与在非灵长类哺乳动物、狐猴和新大陆猴中合成的α - 半乳糖表位结合;以及抗N - 羟乙酰神经氨酸(抗Neu5Gc),它与在猿、旧大陆猴和许多非灵长类哺乳动物中合成的N - 羟乙酰神经氨酸(Neu5Gc)结合。抗Gal在2000万至3000万年前1,3 - 半乳糖基转移酶基因意外失活后出现在旧大陆灵长类动物中。抗Neu5Gc在胞苷 - 单磷酸 - N - 乙酰神经氨酸羟化酶基因失活后出现在人类中,该基因失活导致Neu5Gc在600万年前消失。有人认为,一种致命病毒的流行消灭了合成α - 半乳糖表位的祖先旧大陆灵长类动物,而少数缺乏α - 半乳糖表位并产生抗Gal的突变后代存活了下来,因为抗Gal在亲本群体中复制后会摧毁呈现α - 半乳糖表位的病毒。同样,在致命病毒流行中,抗Neu5Gc保护了少数缺乏Neu5Gc的突变人类,这些流行消灭了合成Neu5Gc的亲本人类。由于α - 半乳糖表位存在于许多人畜共患病病毒上,因此有人认为,提高抗Gal滴度的疫苗在这类人畜共患病病毒流行地区可能具有保护意义。这种保护作用将发生在从非灵长类哺乳动物向人类的病毒传播过程中,但在随后病毒呈现人类聚糖的人传人传播过程中则不会发生。此外,呈现多个α - 半乳糖表位的病毒疫苗的生产会增加其免疫原性,因为抗Gal可有效地将疫苗靶向抗原呈递细胞,使这些细胞大量摄取疫苗。