Stowers Institute for Medical Research, Kansas City, MO, 64110, USA.
Epigenetics Chromatin. 2020 Oct 6;13(1):40. doi: 10.1186/s13072-020-00362-8.
The histone H3K36me3 mark regulates transcription elongation, pre-mRNA splicing, DNA methylation, and DNA damage repair. However, knowledge of the regulation of the enzyme SETD2, which deposits this functionally important mark, is very limited.
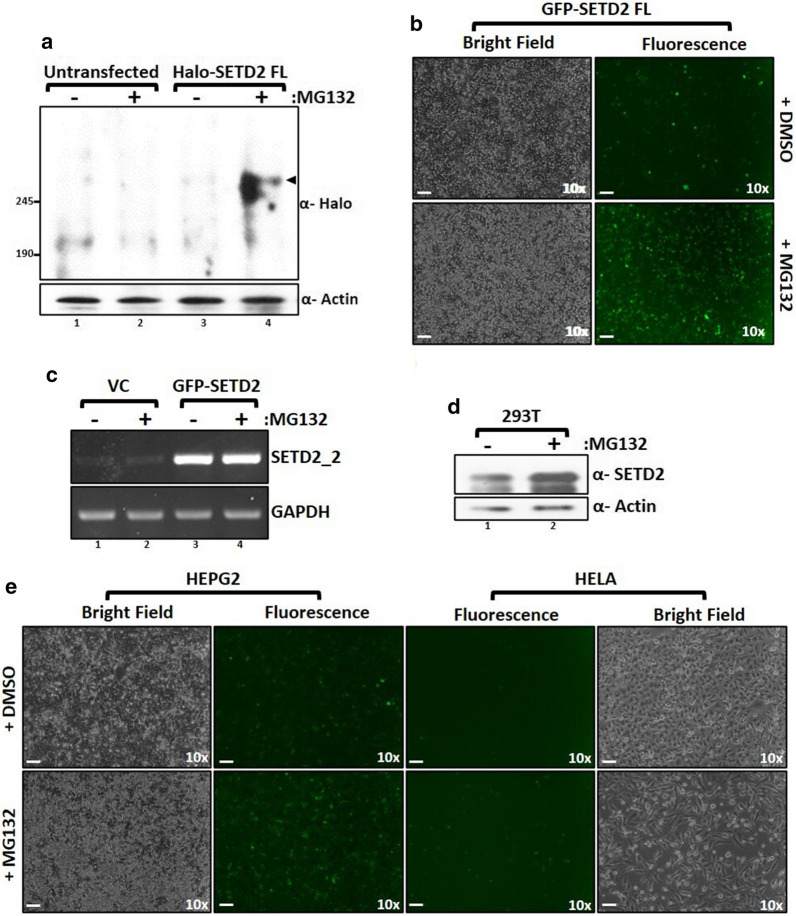
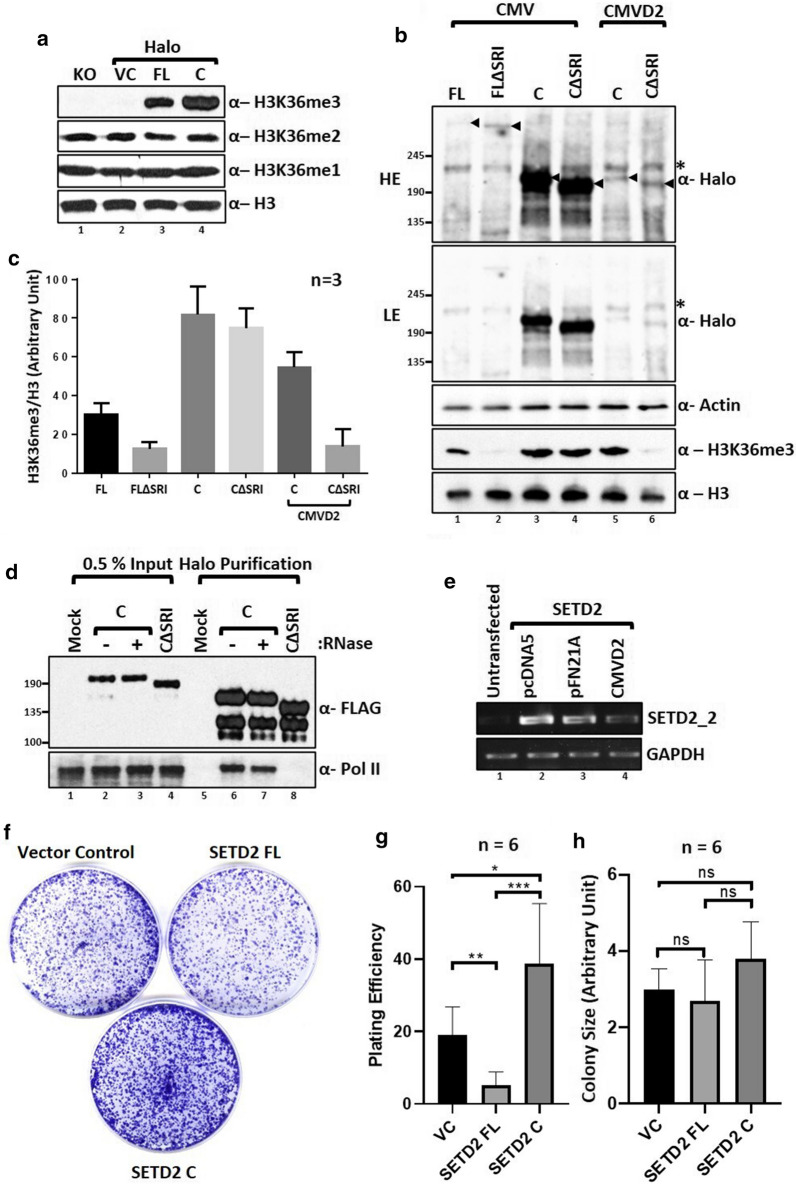
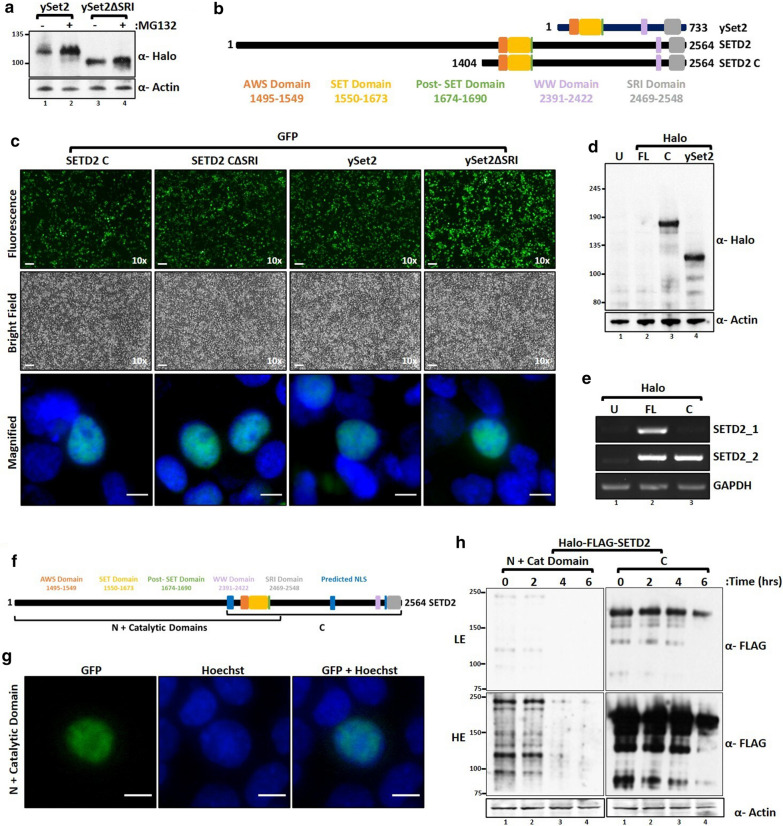
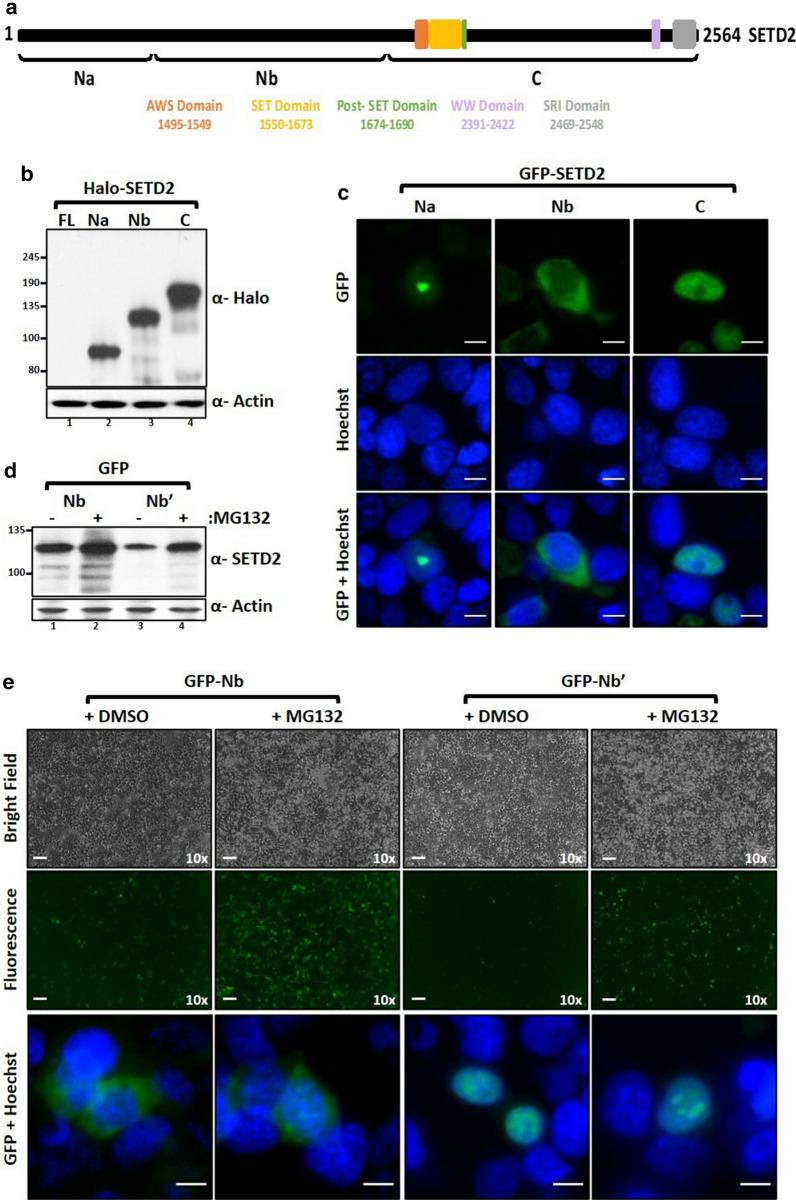
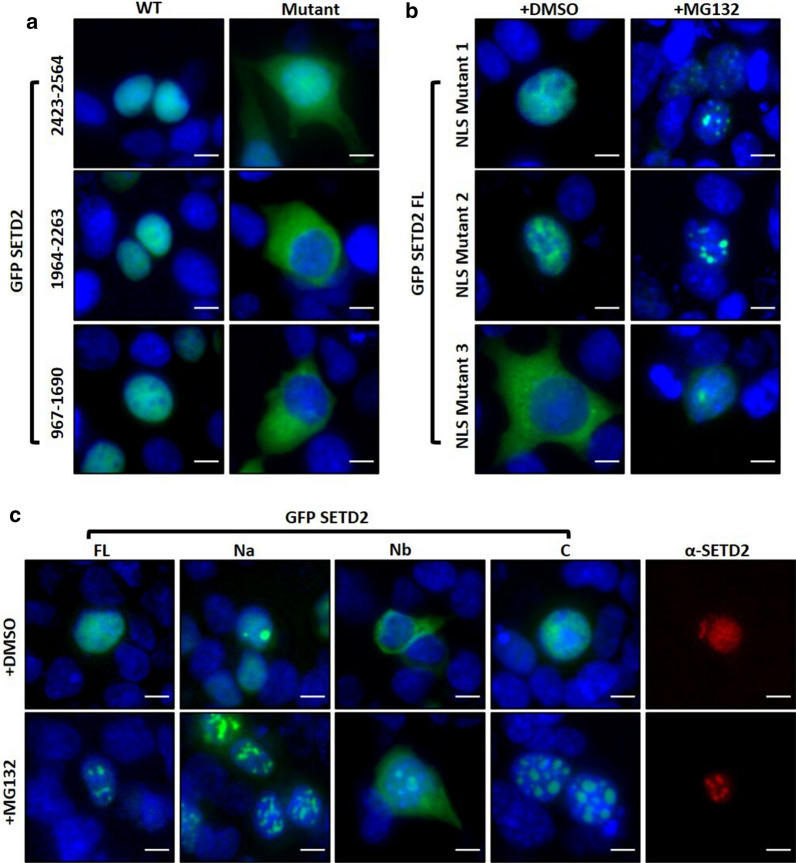
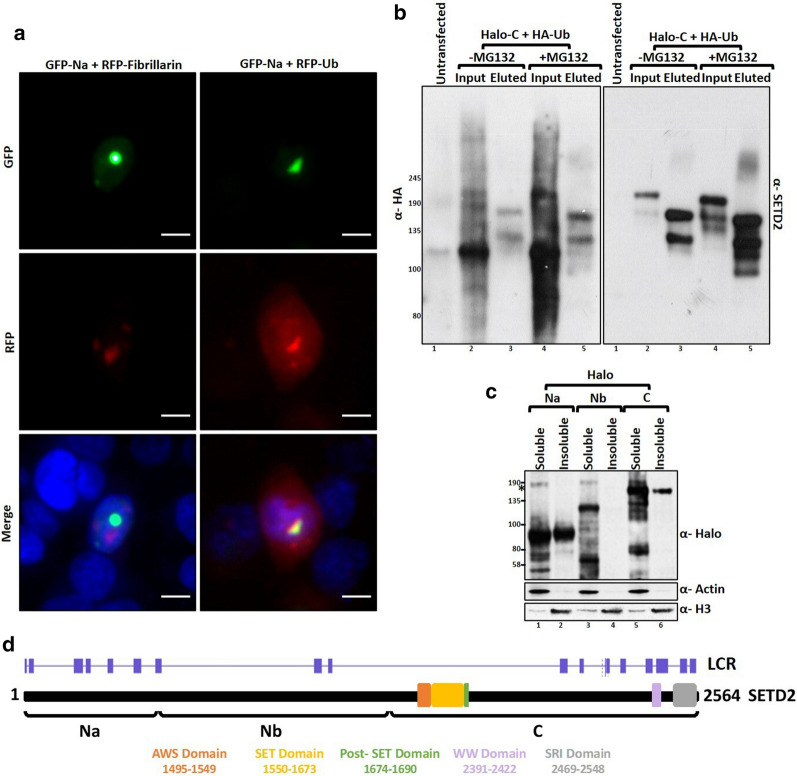
Here, we show that the poorly characterized N-terminal region of SETD2 plays a determining role in regulating the stability of SETD2. This stretch of 1-1403 amino acids contributes to the robust degradation of SETD2 by the proteasome. Besides, the SETD2 protein is aggregate prone and forms insoluble bodies in nuclei especially upon proteasome inhibition. Removal of the N-terminal segment results in the stabilization of SETD2 and leads to a marked increase in global H3K36me3 which, uncharacteristically, happens in a Pol II-independent manner.
The functionally uncharacterized N-terminal segment of SETD2 regulates its half-life to maintain the requisite cellular amount of the protein. The absence of SETD2 proteolysis results in a Pol II-independent H3K36me3 deposition and protein aggregation.
组蛋白 H3K36me3 标记调节转录延伸、前体 mRNA 剪接、DNA 甲基化和 DNA 损伤修复。然而,对于沉积这种具有重要功能标记的酶 SETD2 的调控机制知之甚少。
在这里,我们表明 SETD2 的特征不明显的 N 端区域在调节 SETD2 的稳定性方面起着决定性作用。这 1-1403 个氨基酸的延伸片段有助于 SETD2 被蛋白酶体快速降解。此外,SETD2 蛋白容易聚集,并在核内形成不溶性体,特别是在蛋白酶体抑制时。去除 N 端片段会导致 SETD2 的稳定,并导致全局 H3K36me3 显著增加,这种情况异常地以不依赖 Pol II 的方式发生。
SETD2 功能不明确的 N 端片段调节其半衰期以维持细胞内该蛋白的必需数量。SETD2 蛋白酶解的缺失导致不依赖 Pol II 的 H3K36me3 沉积和蛋白聚集。