Department of Biochemistry & Biophysics, University of California San Francisco, San Francisco, United States.
Department of Pediatrics, Stanford University, Palo Alto, United States.
Elife. 2020 Dec 2;9:e59404. doi: 10.7554/eLife.59404.
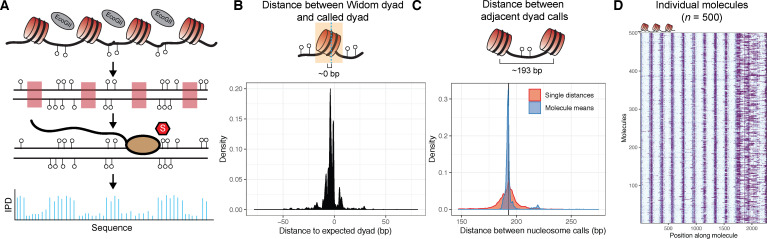
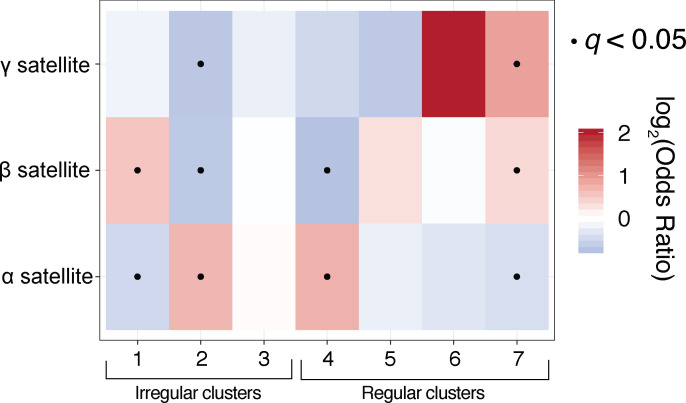
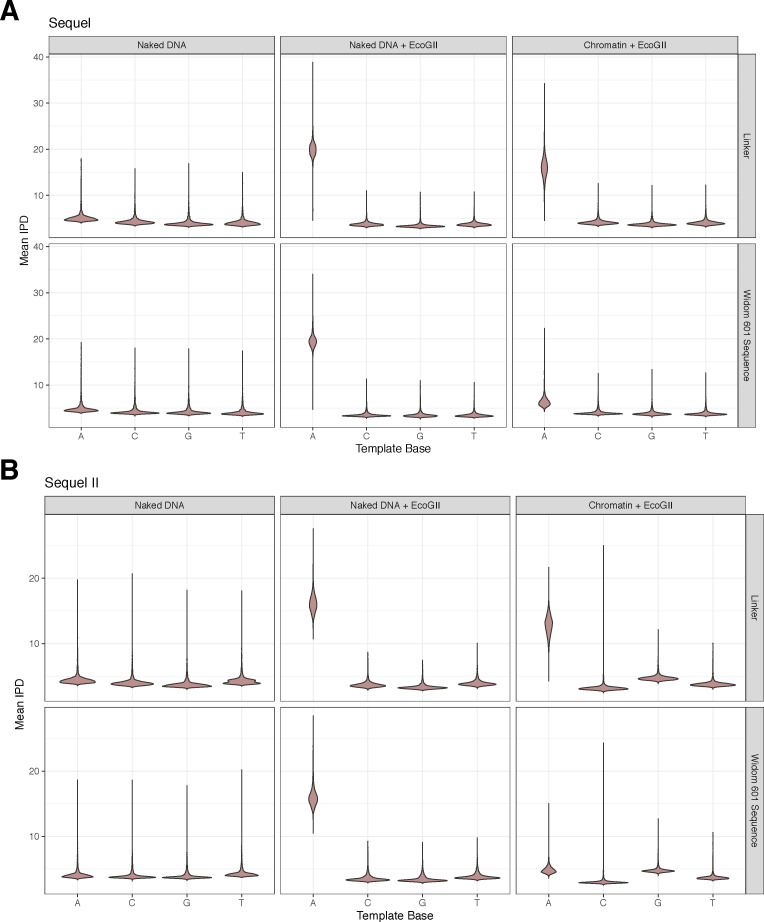
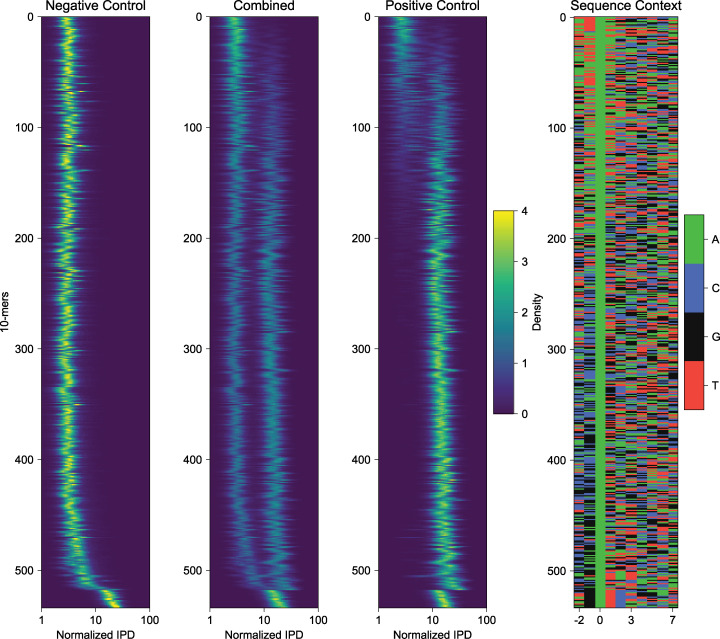
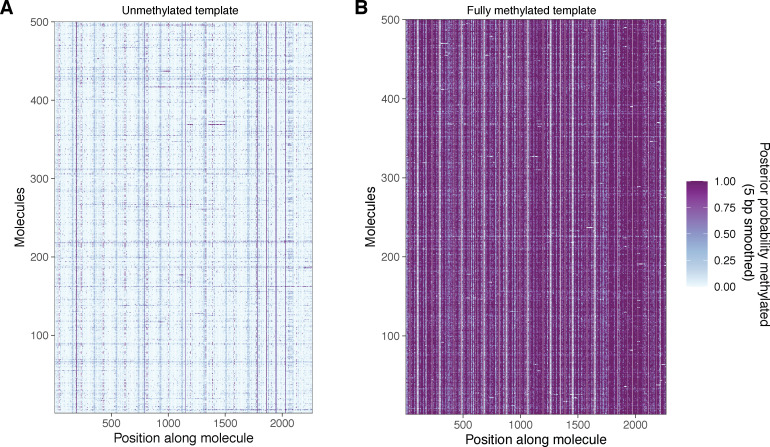
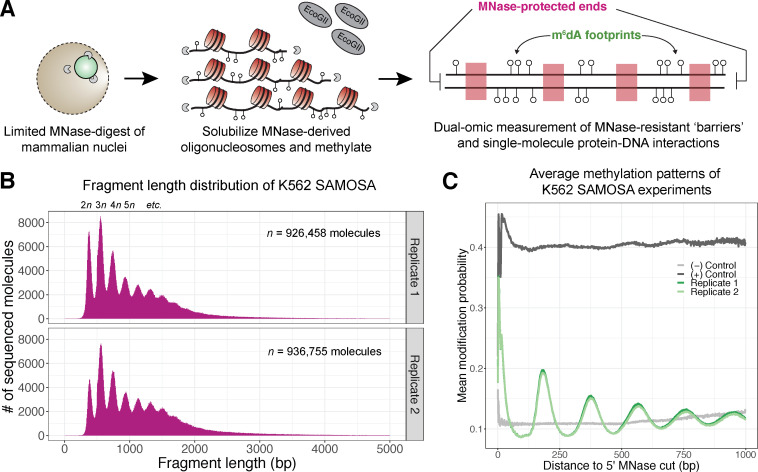
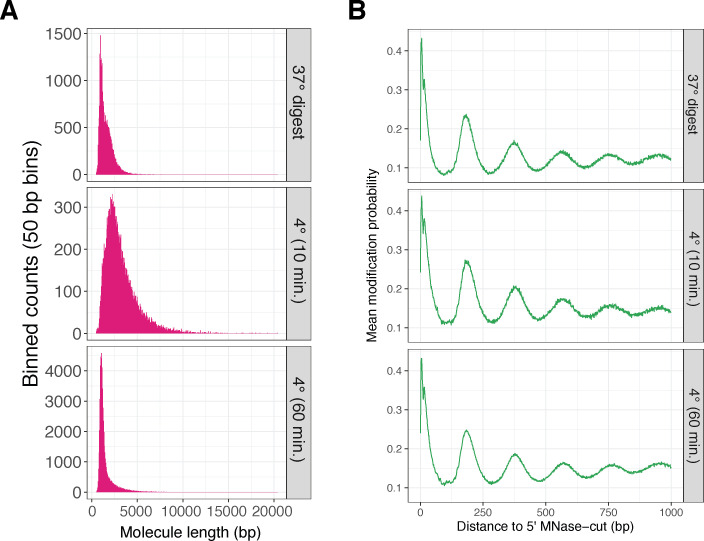
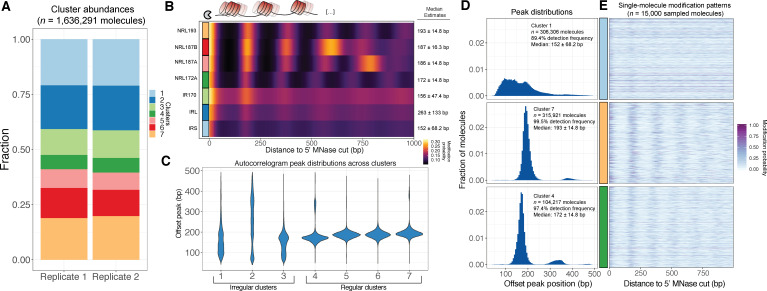
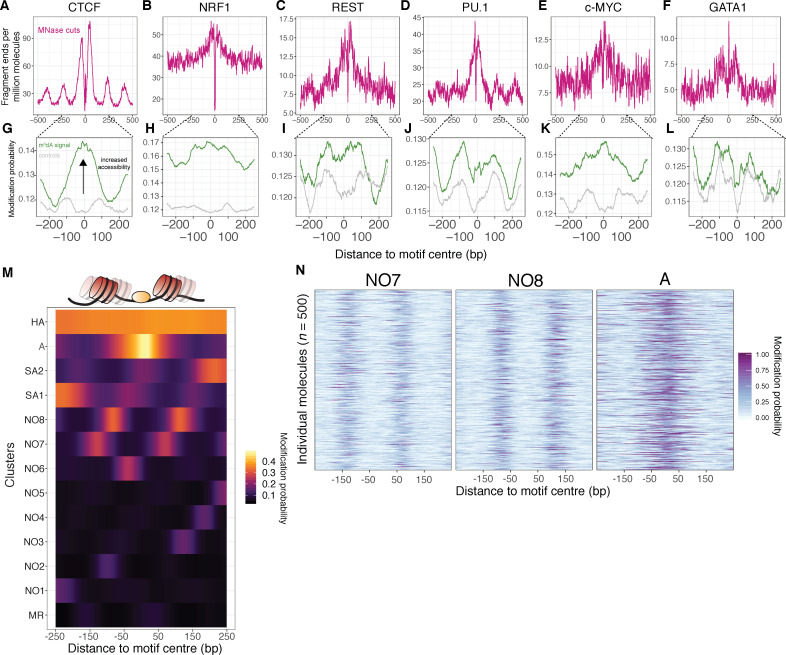
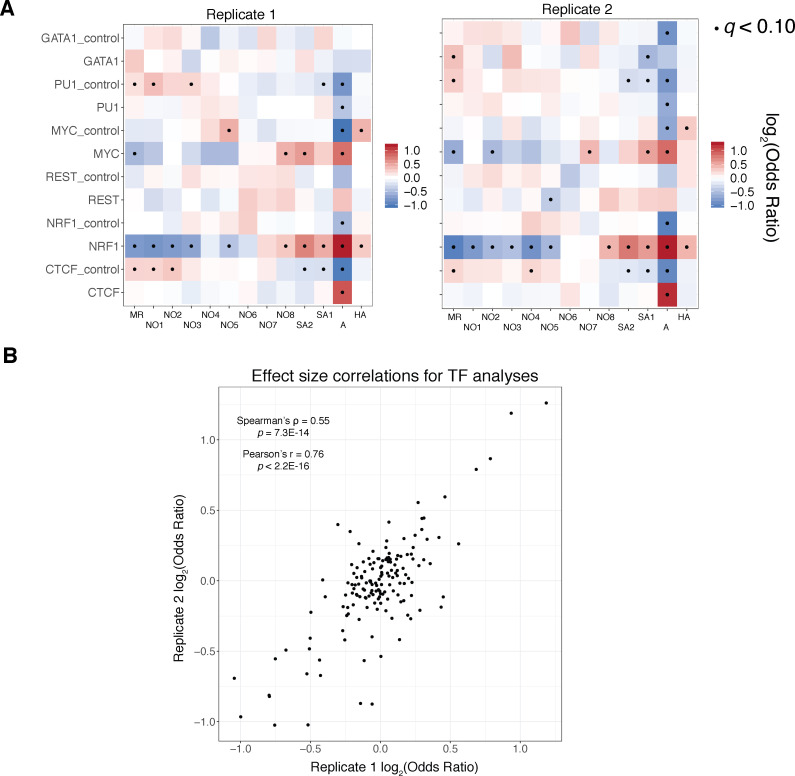
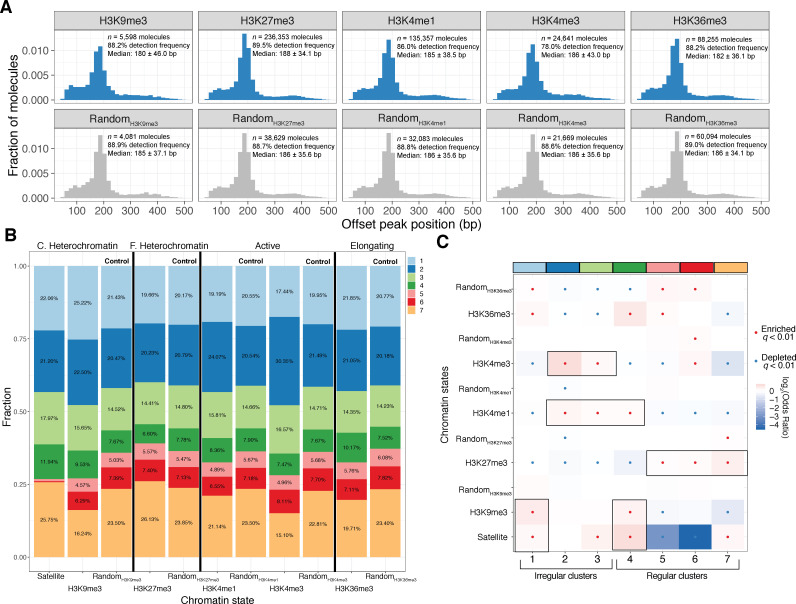
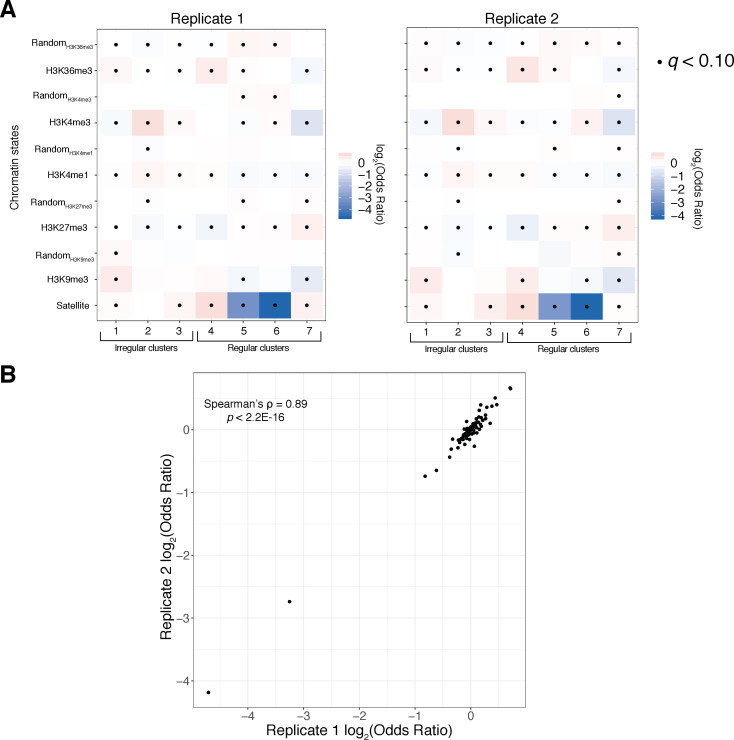
Our understanding of the beads-on-a-string arrangement of nucleosomes has been built largely on high-resolution sequence-agnostic imaging methods and sequence-resolved bulk biochemical techniques. To bridge the divide between these approaches, we present the single-molecule adenine methylated oligonucleosome sequencing assay (SAMOSA). SAMOSA is a high-throughput single-molecule sequencing method that combines adenine methyltransferase footprinting and single-molecule real-time DNA sequencing to natively and nondestructively measure nucleosome positions on individual chromatin fibres. SAMOSA data allows unbiased classification of single-molecular 'states' of nucleosome occupancy on individual chromatin fibres. We leverage this to estimate nucleosome regularity and spacing on single chromatin fibres genome-wide, at predicted transcription factor binding motifs, and across human epigenomic domains. Our analyses suggest that chromatin is comprised of both regular and irregular single-molecular oligonucleosome patterns that differ subtly in their relative abundance across epigenomic domains. This irregularity is particularly striking in constitutive heterochromatin, which has typically been viewed as a conformationally static entity. Our proof-of-concept study provides a powerful new methodology for studying nucleosome organization at a previously intractable resolution and offers up new avenues for modeling and visualizing higher order chromatin structure.
我们对珠串式核小体排列的理解主要建立在高分辨率无序列依赖性成像方法和序列解析的批量生化技术基础上。为了弥合这些方法之间的差距,我们提出了单分子腺嘌呤甲基化寡核小体测序分析(SAMOSA)。SAMOSA 是一种高通量单分子测序方法,它结合了腺嘌呤甲基转移酶足迹法和单分子实时 DNA 测序,以原生和非破坏性的方式测量单个染色质纤维上的核小体位置。SAMOSA 数据允许对单个染色质纤维上核小体占有率的单分子“状态”进行无偏分类。我们利用这一点来估计单个染色质纤维上全基因组、预测的转录因子结合基序和人类表观基因组域中的核小体规则性和间隔。我们的分析表明,染色质由规则和不规则的单分子寡核小体模式组成,它们在整个表观基因组域中的相对丰度略有不同。这种不规则性在组成型异染色质中尤为明显,异染色质通常被视为一种构象静态实体。我们的概念验证研究为在以前难以处理的分辨率下研究核小体组织提供了一种强大的新方法,并为建模和可视化更高阶染色质结构提供了新途径。