Friedrich Miescher Institute for Biomedical Research, CH-4058 Basel, Switzerland.
Faculty of Natural Sciences, University of Basel, CH-4056 Basel, Switzerland.
Genes Dev. 2021 Jan 1;35(1-2):82-101. doi: 10.1101/gad.344234.120. Epub 2020 Dec 10.
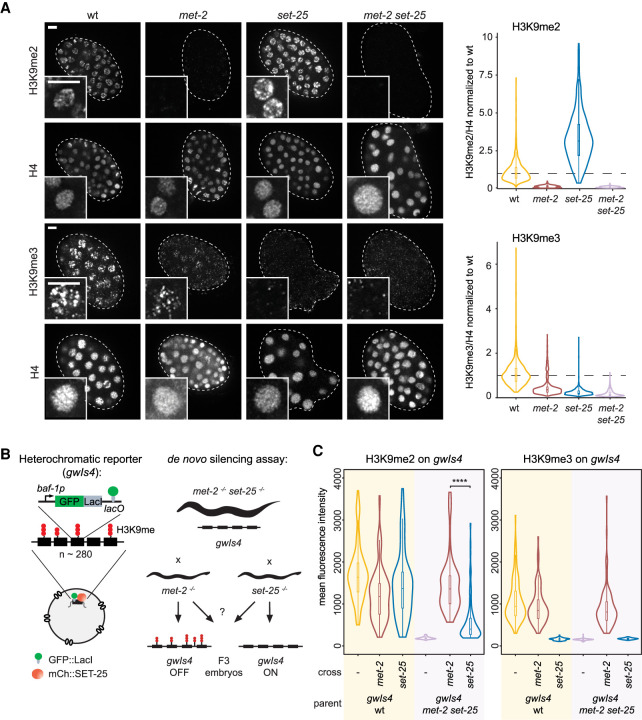
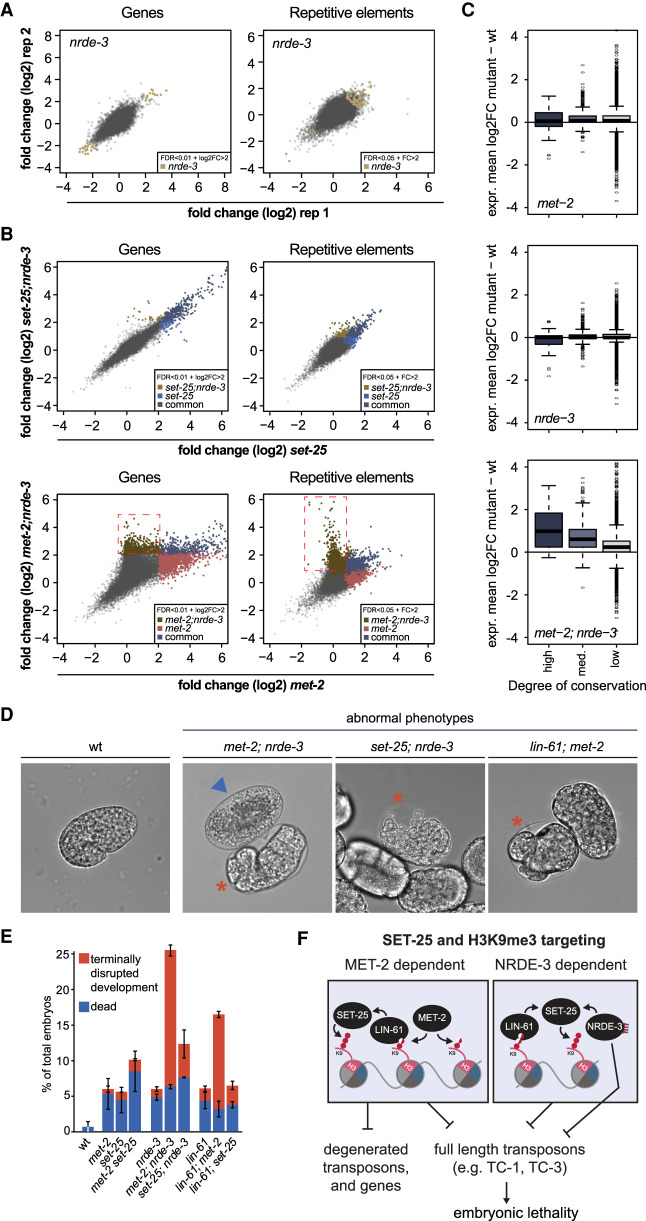
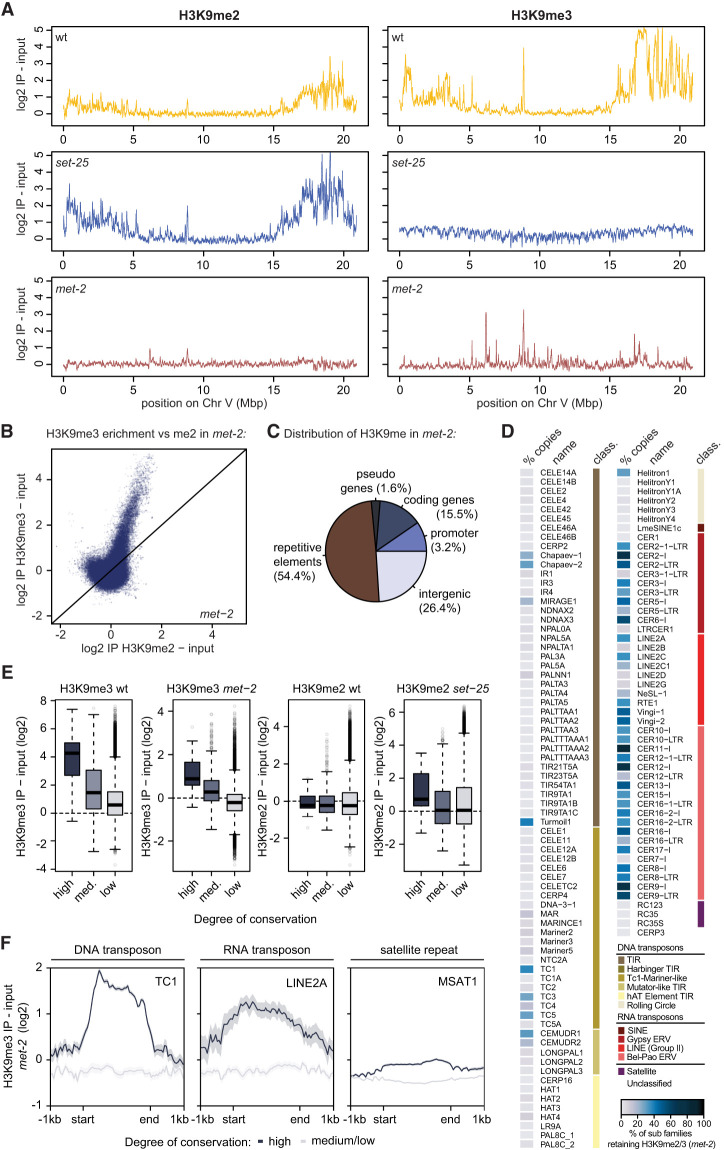
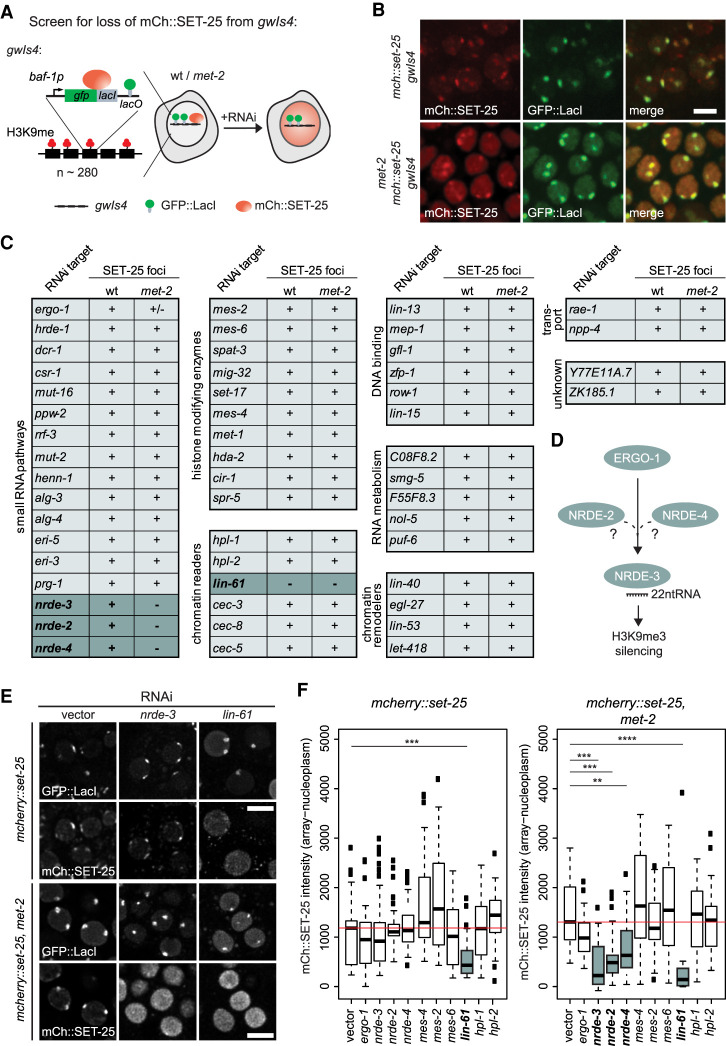
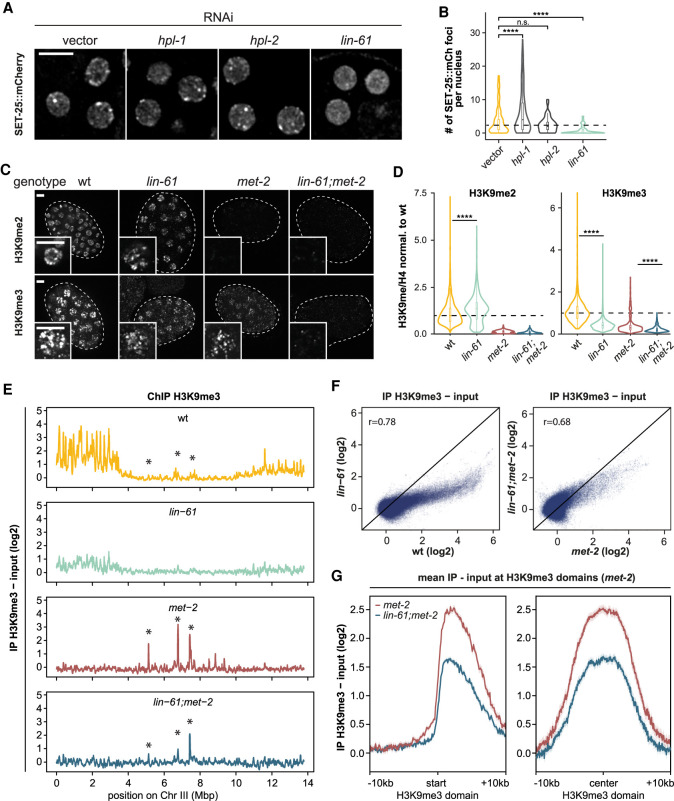
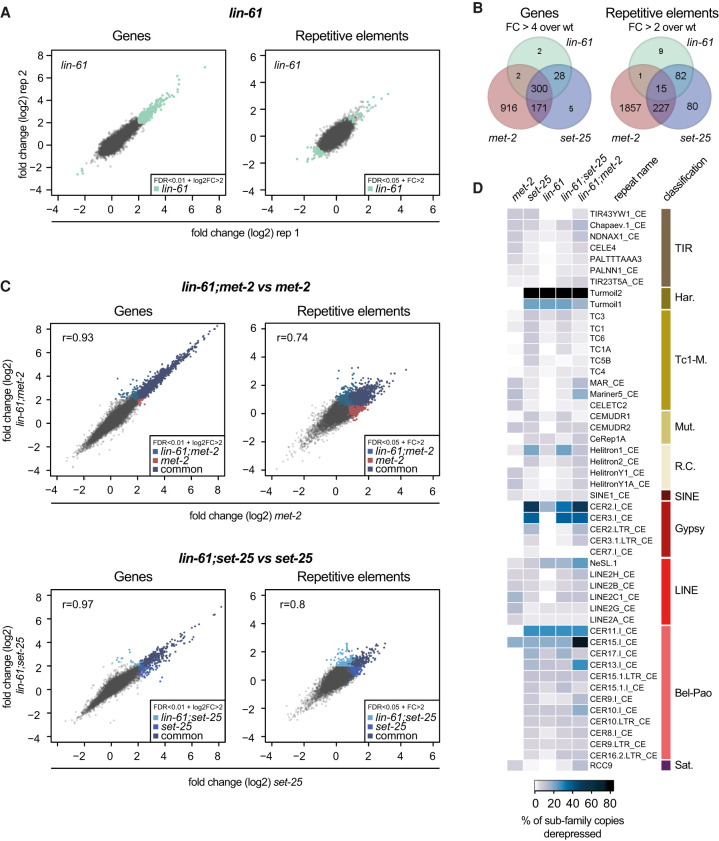
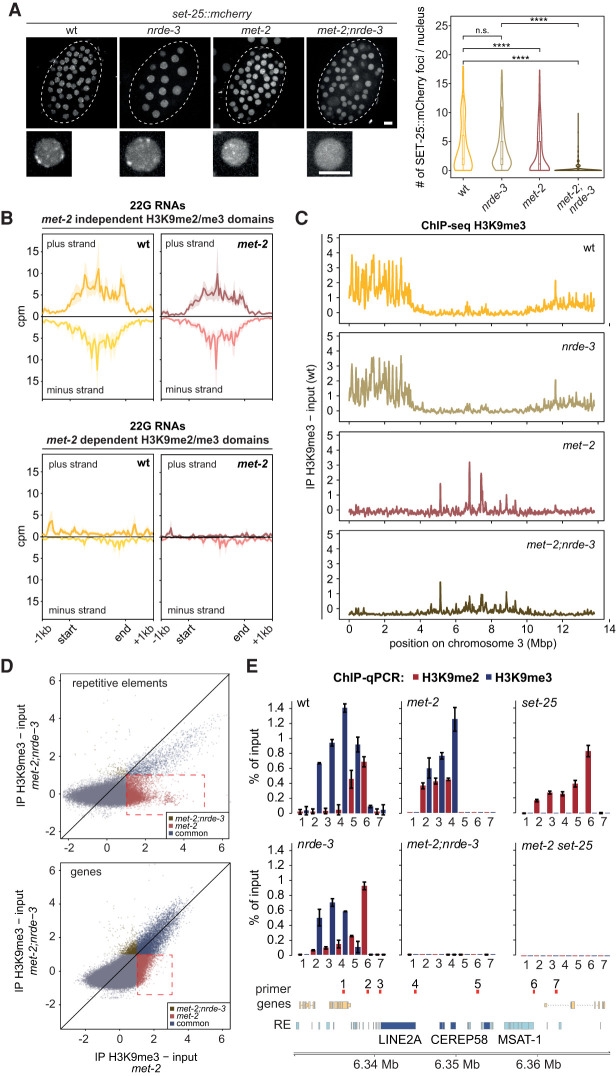
The establishment and maintenance of chromatin domains shape the epigenetic memory of a cell, with the methylation of histone H3 lysine 9 (H3K9me) defining transcriptionally silent heterochromatin. We show here that the SET-25 (SUV39/G9a) histone methyltransferase (HMT), which catalyzes H3K9me1, me2 and me3, can establish repressed chromatin domains de novo unlike the SETDB1 homolog MET-2. Thus, SET-25 is needed to silence novel insertions of RNA or DNA transposons, and repress tissue-specific genes de novo during development. We identify two partially redundant pathways that recruit SET-25 to its targets. One pathway requires LIN-61 (L3MBTL2), which uses its four MBT domains to bind the H3K9me2 deposited by MET-2. The second pathway functions independently of MET-2 and involves the somatic Argonaute NRDE-3 and small RNAs. This pathway targets primarily highly conserved RNA and DNA transposons. These redundant SET-25 targeting pathways (MET-2-LIN-61-SET-25 and NRDE-3-SET-25) ensure repression of intact transposons and de novo insertions, while MET-2 can act alone to repress simple and satellite repeats. Removal of both pathways in the double mutant leads to the loss of somatic H3K9me2 and me3 and the synergistic derepression of transposons in embryos, strongly elevating embryonic lethality.
染色质域的建立和维持塑造了细胞的表观遗传记忆,组蛋白 H3 赖氨酸 9(H3K9me)的甲基化定义了转录沉默的异染色质。我们在这里表明,SET-25(SUV39/G9a)组蛋白甲基转移酶(HMT),它催化 H3K9me1、me2 和 me3,可以从头建立被抑制的染色质域,而 SETDB1 同源物 MET-2 则不能。因此,SET-25 对于沉默 RNA 或 DNA 转座子的新插入以及在发育过程中从头抑制组织特异性基因是必需的。我们确定了两种部分冗余的途径,这些途径将 SET-25 招募到其靶标上。一种途径需要 LIN-61(L3MBTL2),它使用其四个 MBT 结构域结合 MET-2 沉积的 H3K9me2。第二种途径独立于 MET-2 起作用,涉及体细胞 Argonaute NRDE-3 和小 RNA。该途径主要靶向高度保守的 RNA 和 DNA 转座子。这些冗余的 SET-25 靶向途径(MET-2-LIN-61-SET-25 和 NRDE-3-SET-25)确保了完整转座子和新插入的抑制,而 MET-2 可以单独起作用抑制简单和卫星重复序列。在双突变体中去除这两种途径会导致体细胞 H3K9me2 和 me3 的丧失以及转座子的协同去抑制,强烈提高胚胎的致死率。