Friedrich Miescher Institute for Biomedical Research, Basel, Switzerland.
Friedrich Miescher Institute for Biomedical Research, Basel, Switzerland
J Cell Biol. 2019 Mar 4;218(3):820-838. doi: 10.1083/jcb.201811038. Epub 2019 Feb 8.
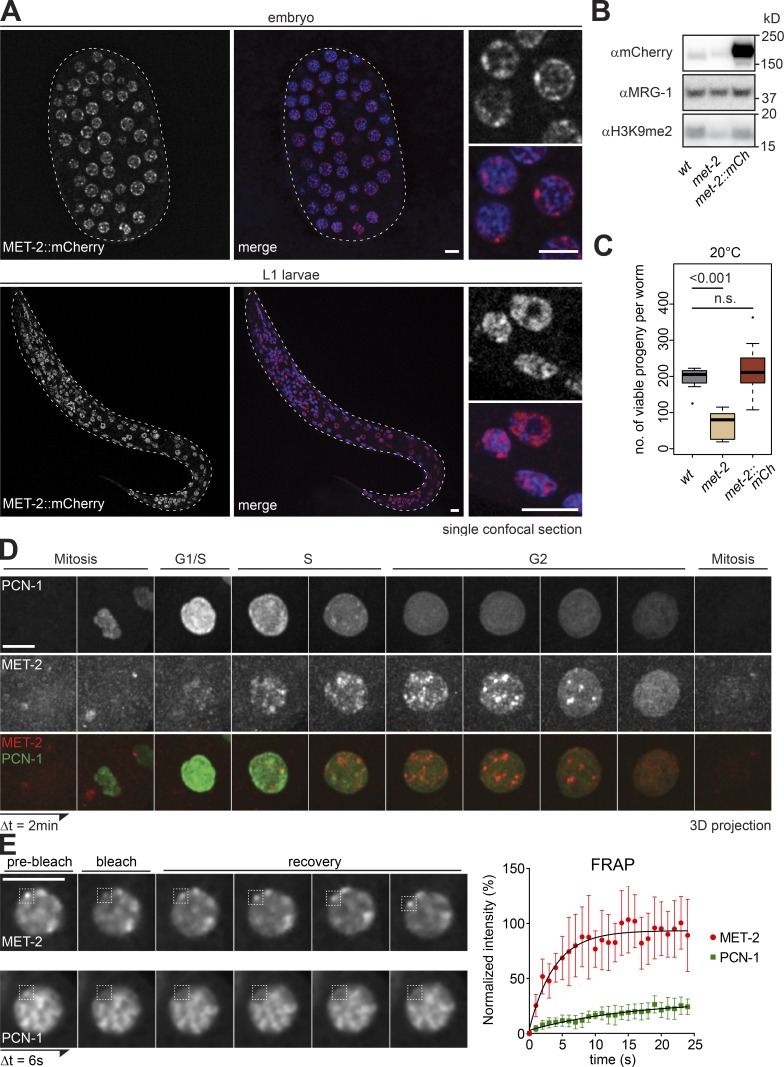
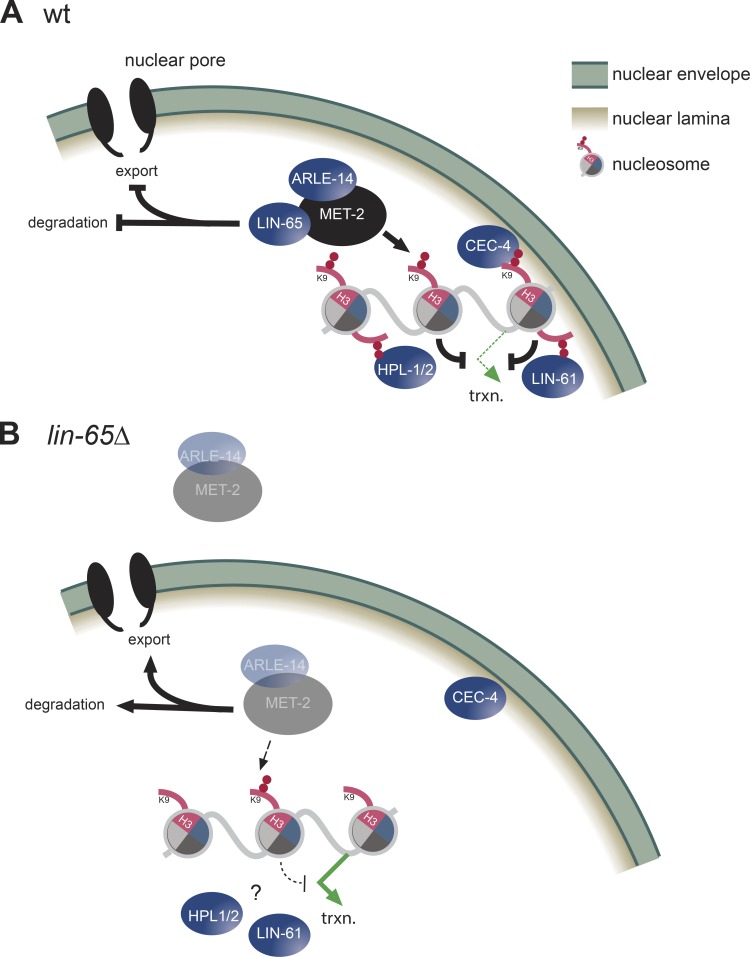
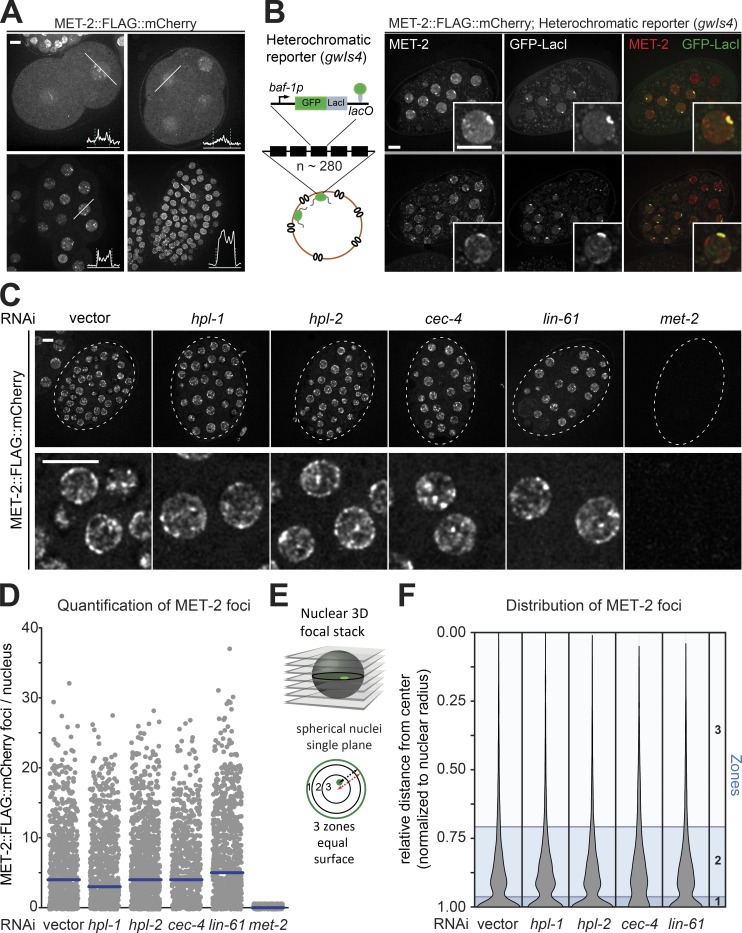
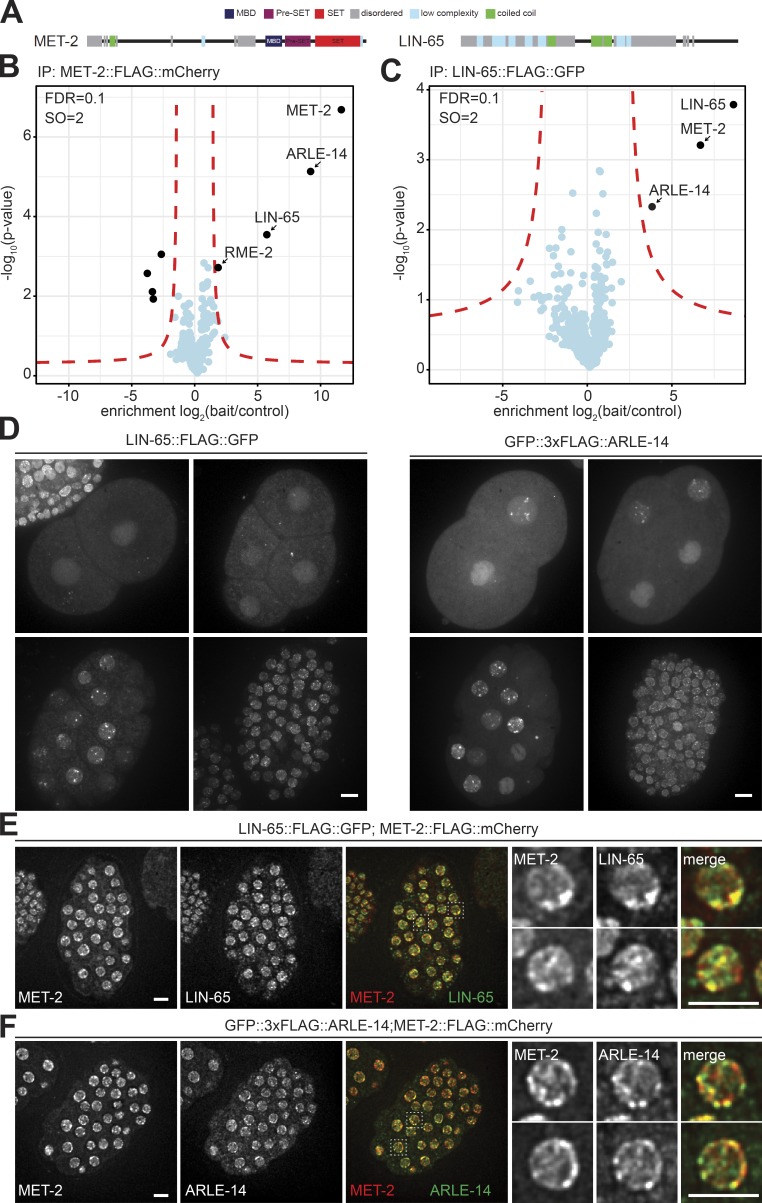
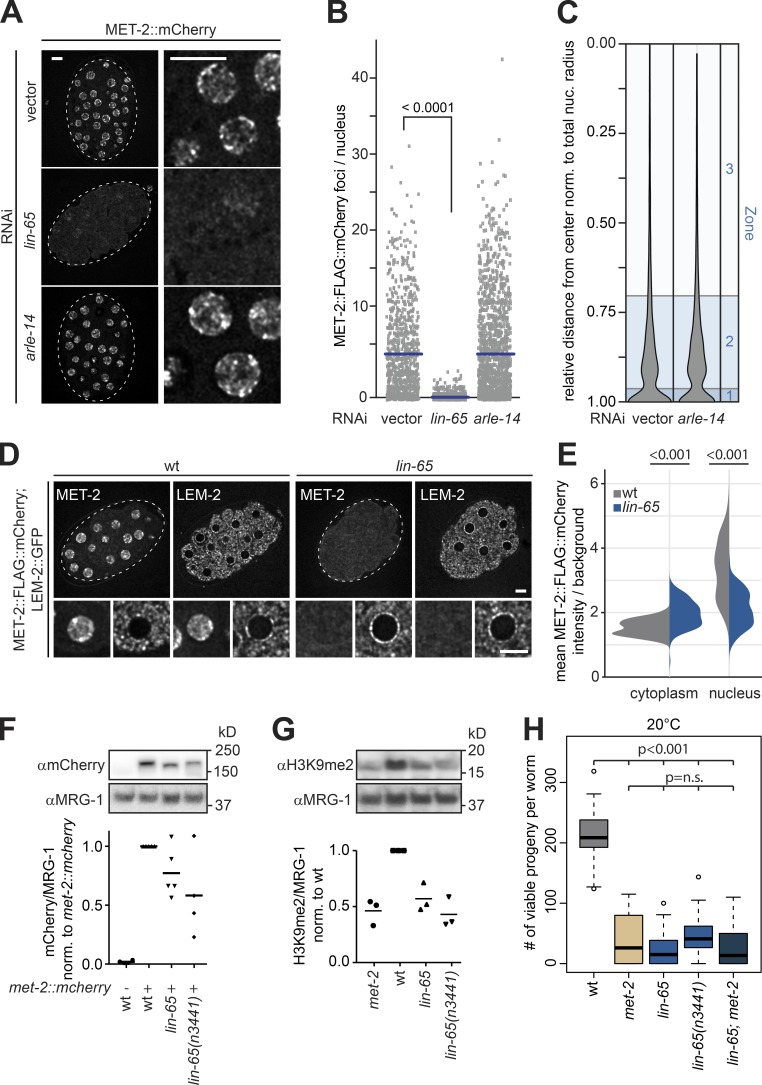
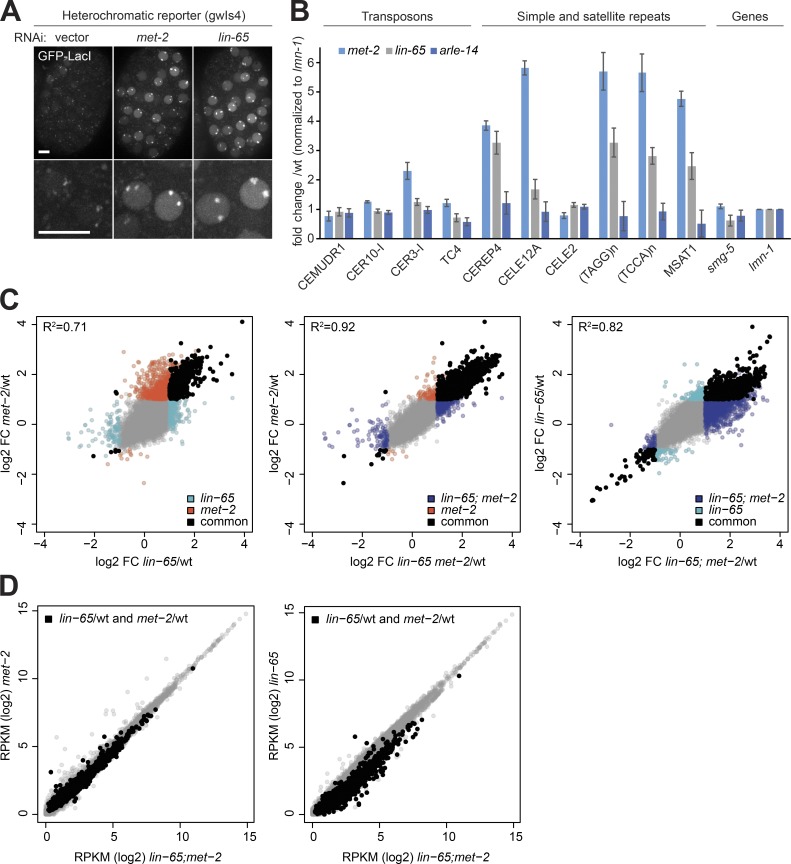
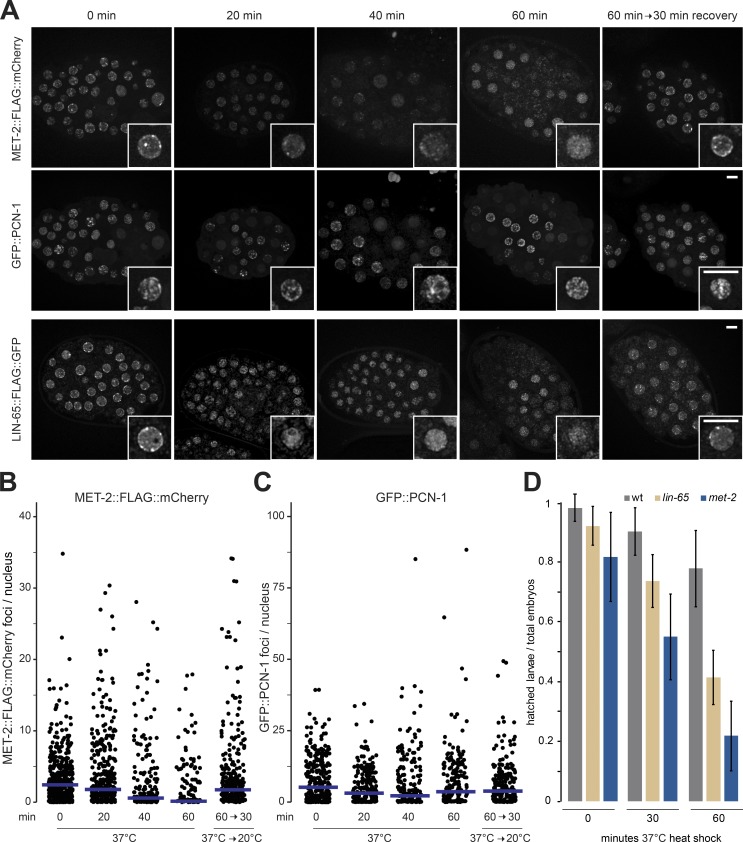
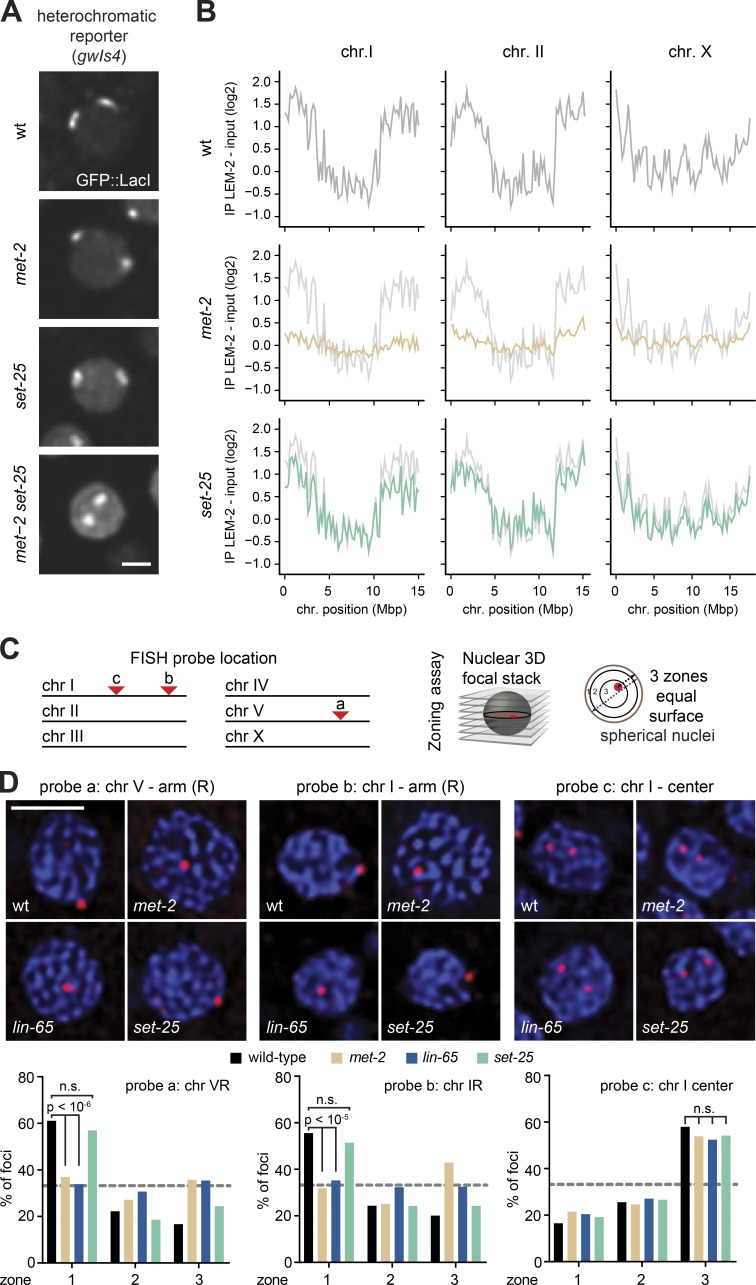
The segregation of the genome into accessible euchromatin and histone H3K9-methylated heterochromatin helps silence repetitive elements and tissue-specific genes. In , MET-2, the homologue of mammalian SETDB1, catalyzes H3K9me1 and me2, yet like SETDB1, its regulation is enigmatic. Contrary to the cytosolic enrichment of overexpressed MET-2, we show that endogenous MET-2 is nuclear throughout development, forming perinuclear foci in a cell cycle-dependent manner. Mass spectrometry identified two cofactors that bind MET-2: LIN-65, a highly unstructured protein, and ARLE-14, a conserved GTPase effector. All three factors colocalize in heterochromatic foci. Ablation of , but not , mislocalizes and destabilizes MET-2, resulting in decreased H3K9 dimethylation, dispersion of heterochromatic foci, and derepression of MET-2 targets. Mutation of or also disrupts the perinuclear anchoring of genomic heterochromatin. Loss of LIN-65, like that of MET-2, compromises temperature stress resistance and germline integrity, which are both linked to promiscuous repeat transcription and gene expression.
基因组的区隔成可及的常染色质和组蛋白 H3K9 甲基化异染色质有助于沉默重复元件和组织特异性基因。在, MET-2,其同源物的哺乳动物 SETDB1,催化 H3K9me1 和 me2,然而,像 SETDB1,其调节是神秘的。与细胞质中富含过表达的 MET-2 相反,我们发现内源性 MET-2 在整个发育过程中都是核的,以细胞周期依赖性的方式形成核周焦点。质谱分析鉴定了两个与 MET-2 结合的辅助因子:LIN-65,一种高度非结构化的蛋白质,和 ARLE-14,一种保守的 GTPase 效应因子。这三个因素都在异染色质焦点中共定位。, 而不是, 的缺失导致 MET-2 的定位错误和不稳定性,导致 H3K9 二甲基化减少、异染色质焦点分散和 MET-2 靶基因的去抑制。 或 突变也破坏了基因组异染色质的核周锚定。LIN-65 的缺失,就像 MET-2 的缺失一样,会影响温度应激抗性和生殖系完整性,这两者都与随意重复转录和基因表达有关。