The Laboratory of Cell Biochemistry and Topogenetic Regulation, College of Bioengineering and Faculty of Medical Sciences, Chongqing University, No. 174 Shazheng Street, Shapingba District, Chongqing 400044, China.
Department of Biochemistry and Molecular Biology, College of Basic Medical Sciences, Southwest Medical University, Sichuan 646000, China.
Oxid Med Cell Longev. 2020 Nov 16;2020:5097109. doi: 10.1155/2020/5097109. eCollection 2020.
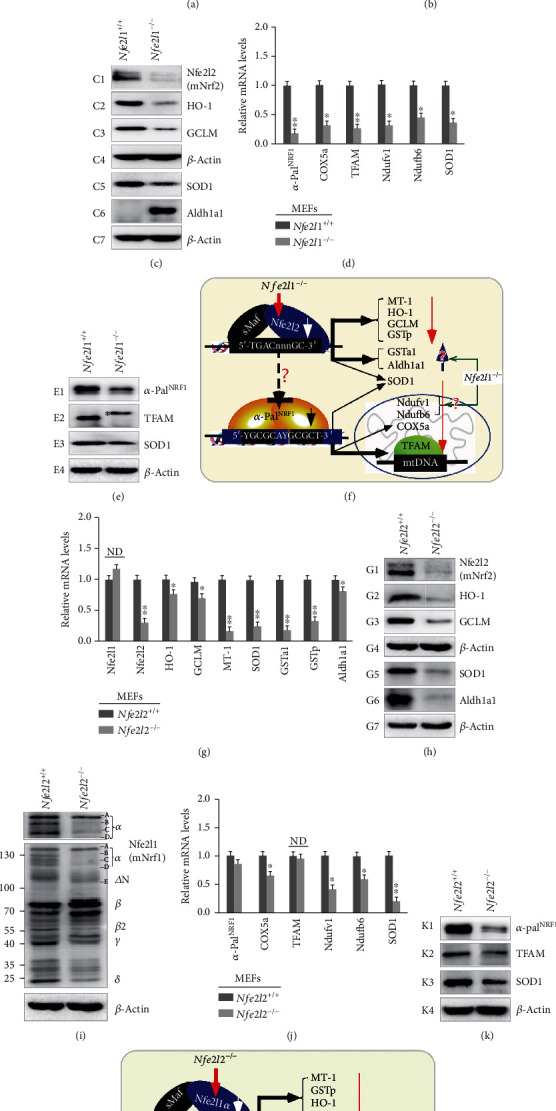
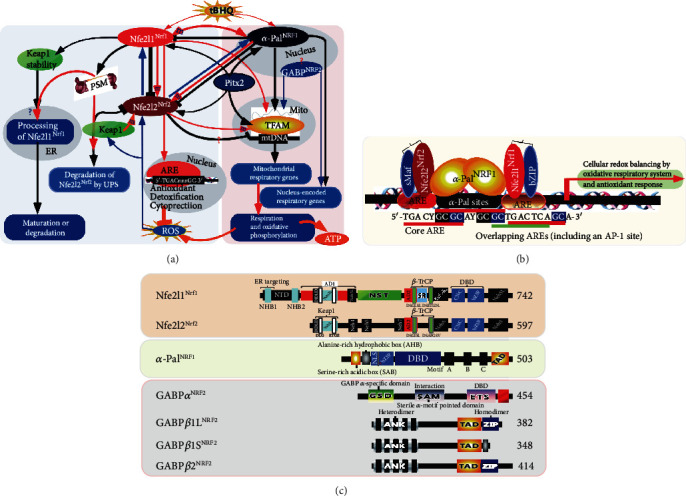
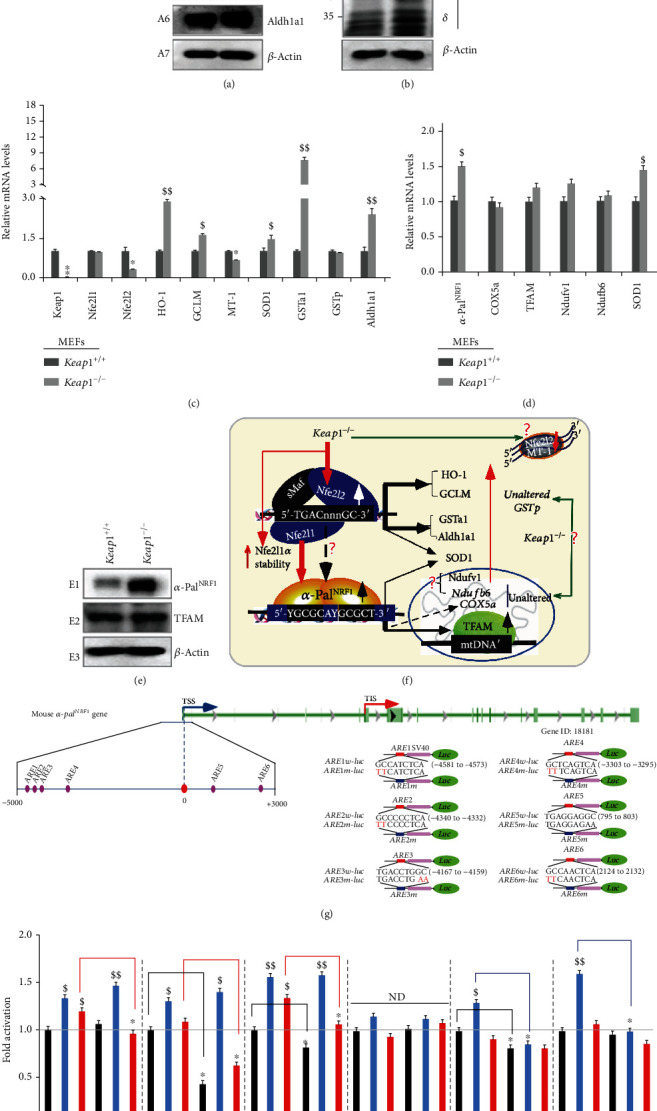
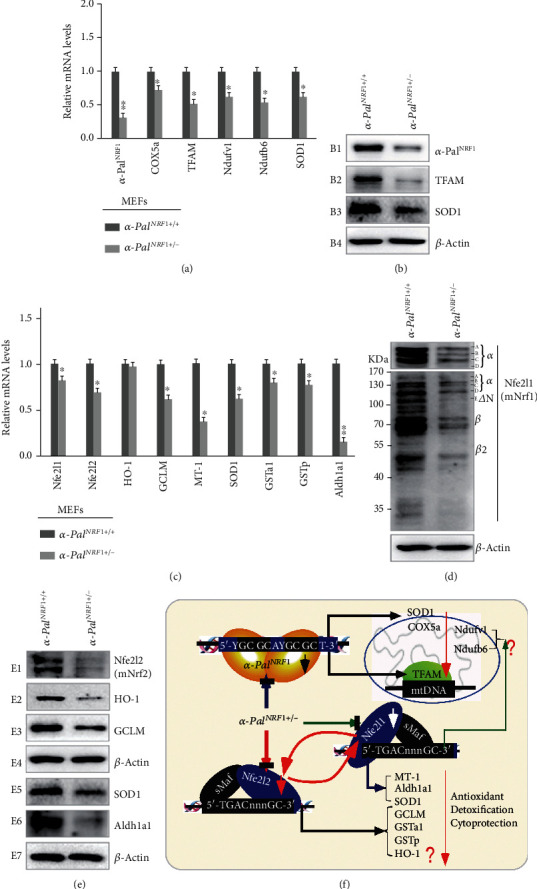
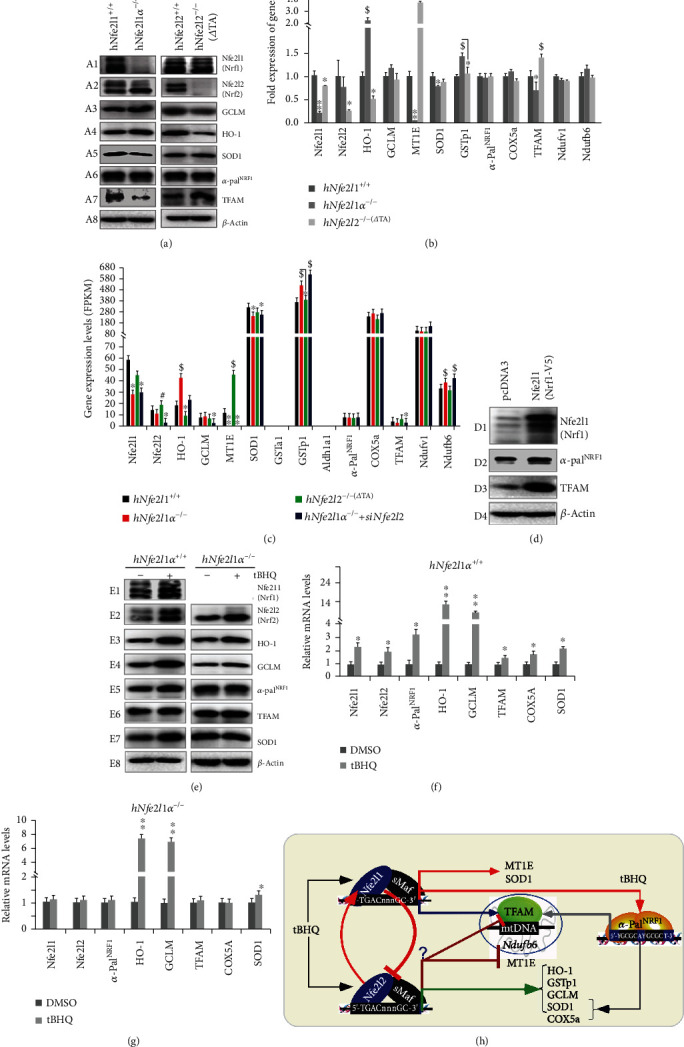
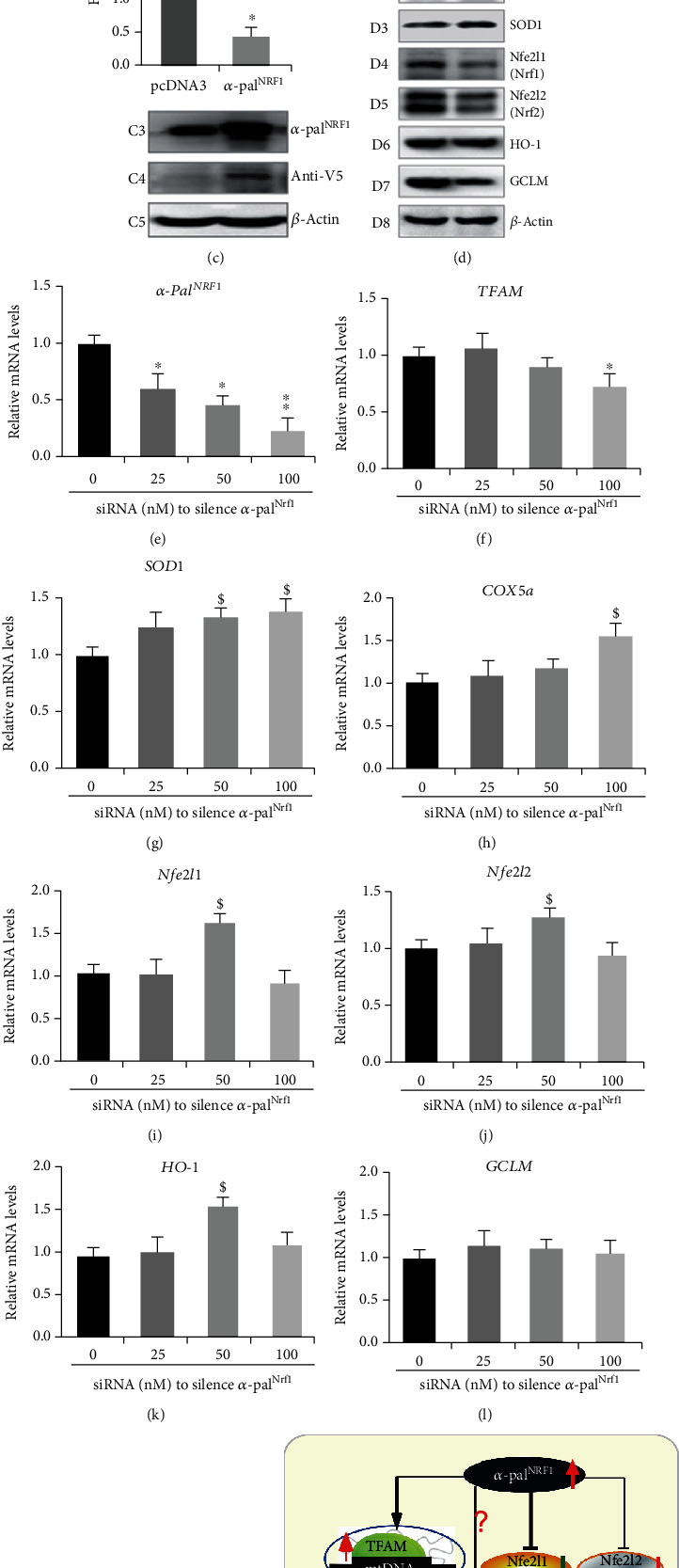
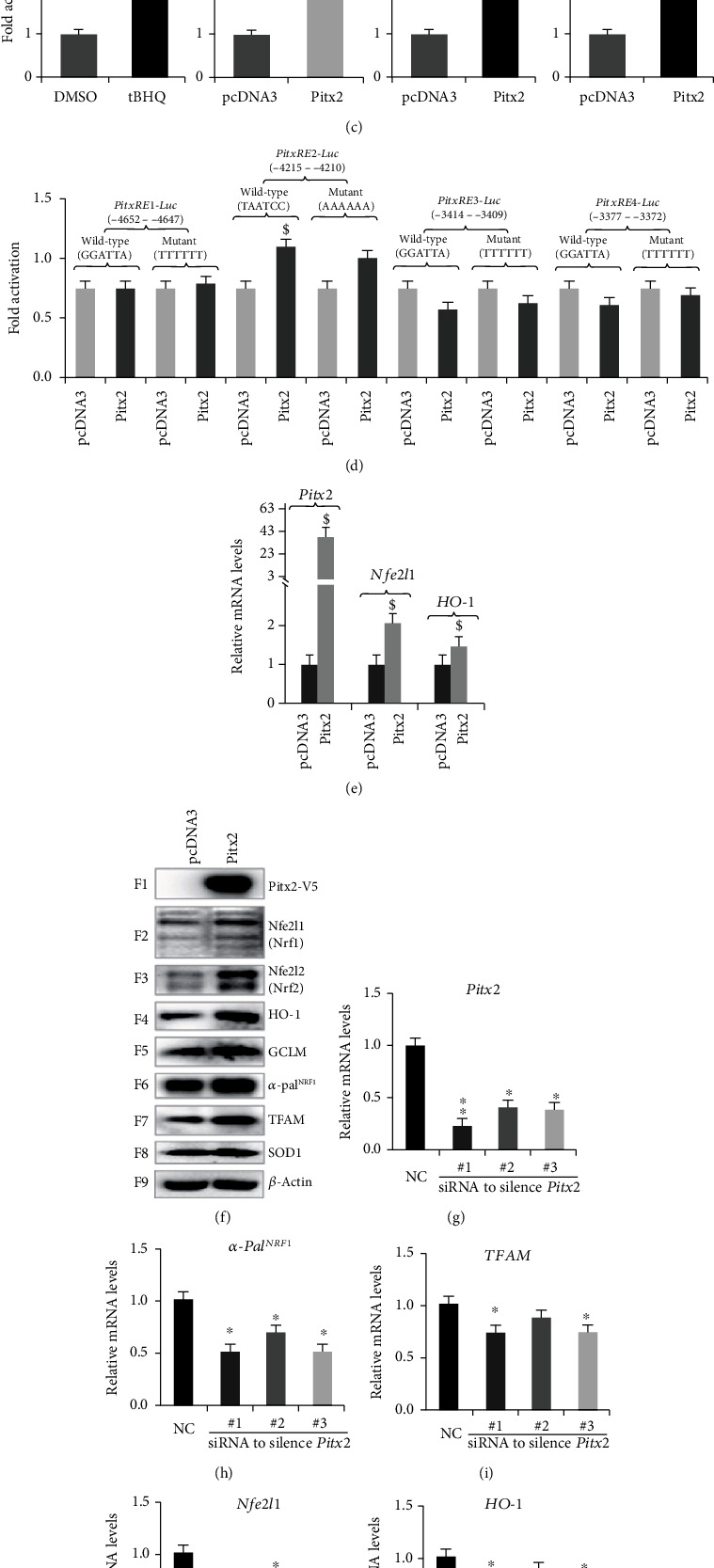
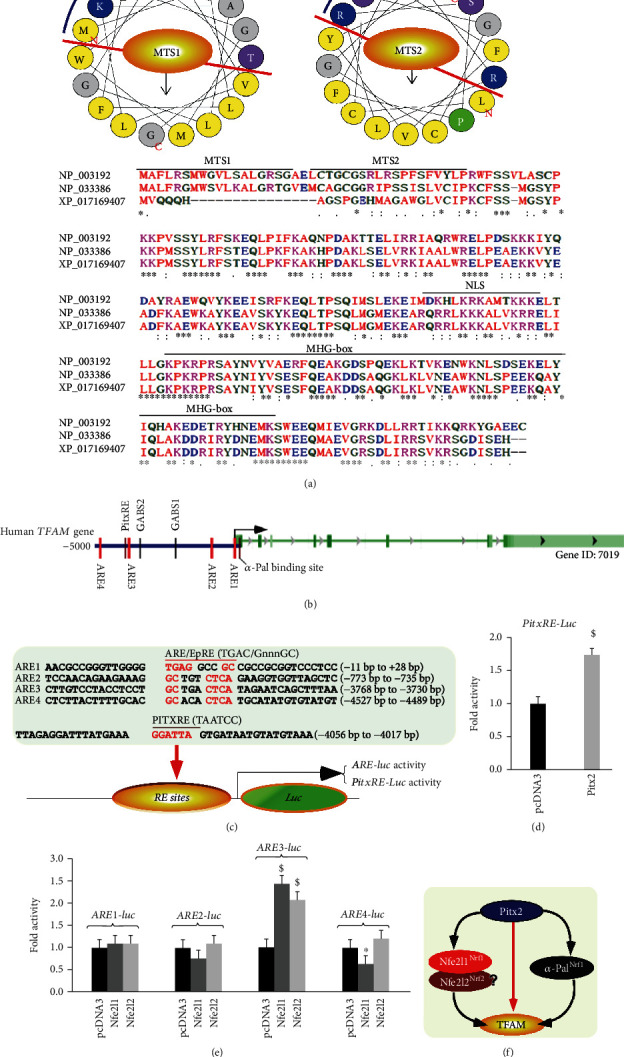
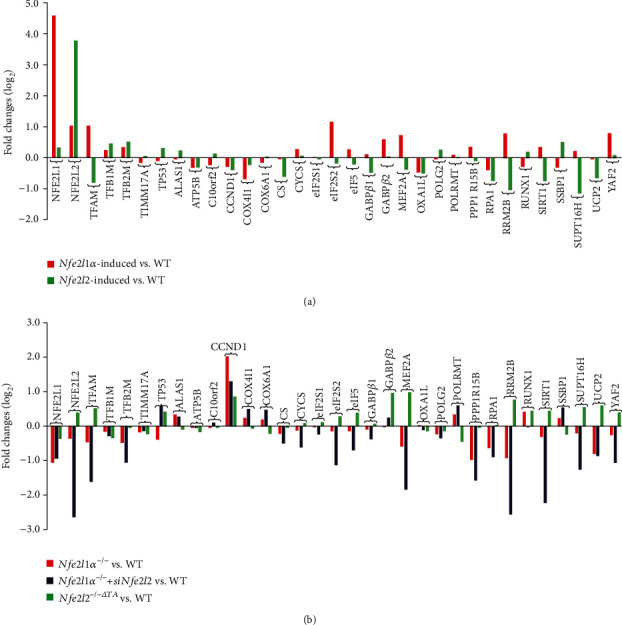
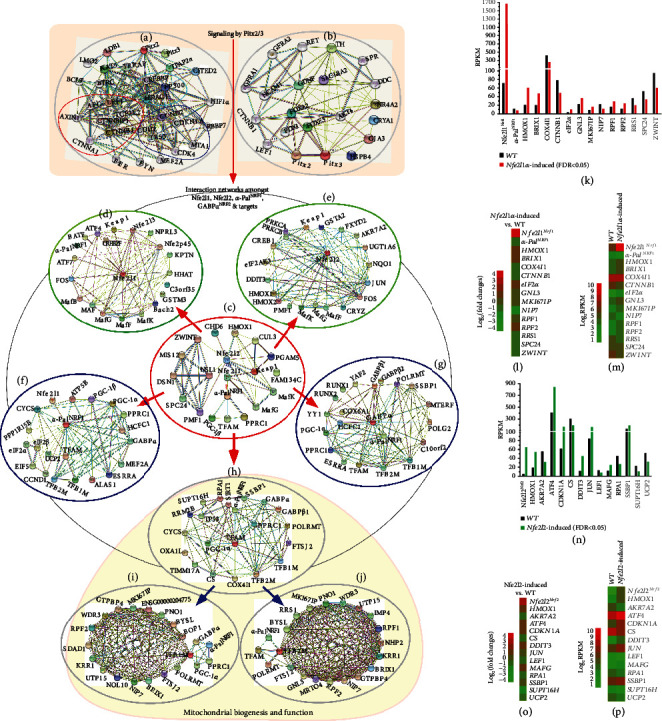
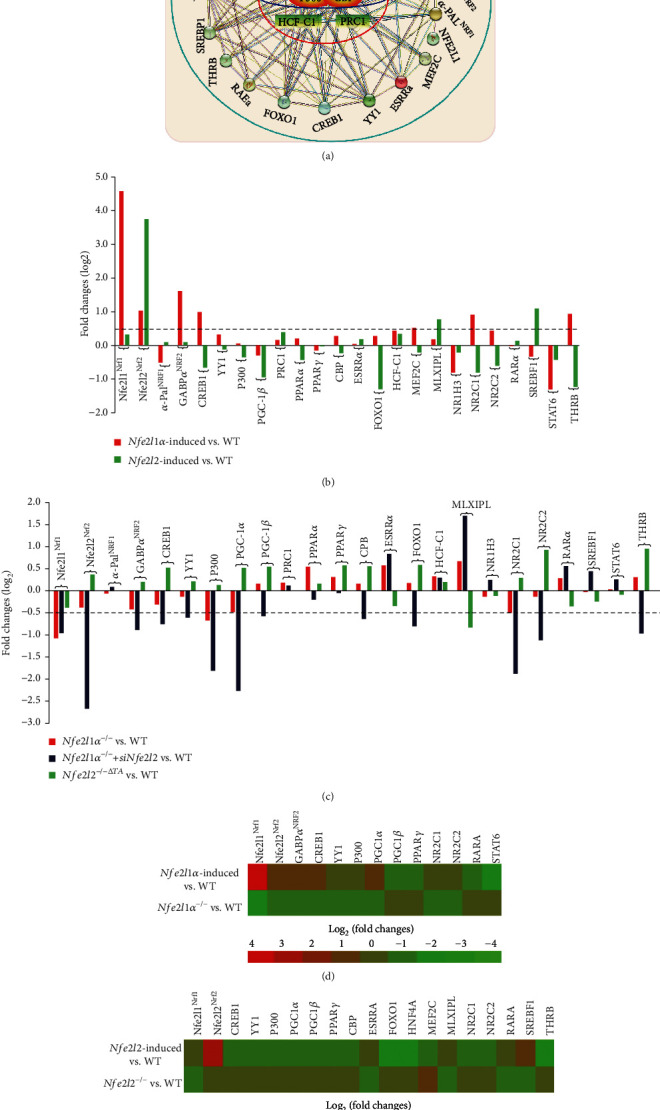
There is hitherto no literature available for explaining two distinct, but confused, Nrf1 transcription factors, because they shared the same abbreviations from nuclear factor erythroid 2-related factor 1 (also called Nfe2l1) and nuclear respiratory factor (originally designated -Pal). Thus, we have here identified that Nfe2l1 and -Pal exert synergistic and antagonistic roles in integrative regulation of the nuclear-to-mitochondrial respiratory and antioxidant transcription profiles. In mouse embryonic fibroblasts (MEFs), knockout of leads to substantial decreases in expression levels of -Pal and Nfe2l2, together with TFAM (mitochondrial transcription factor A) and other target genes. Similar inhibitory results were determined in MEFs but with an exception that both and were distinguishably upregulated in MEFs. Such synergistic contributions of Nfe2l1 and Nfe2l2 to the positive regulation of -Pal and TFAM were validated in MEFs. However, human -Pal expression was unaltered by , , or even , albeit TFAM was activated by Nfe2l1 but inhibited by Nfe2l2; such an antagonism occurred in HepG2 cells. Conversely, almost all of mouse Nfe2l1, Nfe2l2, and cotarget genes were downexpressed in MEFs. On the contrary, upregulation of human Nfe2l1, Nfe2l2, and relevant reporter genes took place after silencing of -Pal, but their downregulation occurred upon ectopic expression of -Pal. Furtherly, Pitx2 (pituitary homeobox 2) was also identified as a direct upstream regulator of Nfe2l1 and TFAM, besides -Pal. Overall, these across-talks amongst Nfe2l1, Nfe2l2, and -Pal, along with Pitx2, are integrated from the endoplasmic reticulum towards the nuclear-to-mitochondrial communication for targeting TFAM, in order to finely tune the robust balance of distinct cellular oxidative respiratory and antioxidant gene transcription networks, albeit they differ between the mouse and the human. In addition, it is of crucial importance to note that, in view of such mutual interregulation of these transcription factors, much cautions should be severely taken for us to interpret those relevant experimental results obtained from knockout of Nfe2l1, Nfe2l2, -Pal or Pitx2, or their gain-of-functional mutants.
目前尚无文献能够解释两种截然不同但又混淆不清的 Nrf1 转录因子,因为它们共享相同的缩写词,即核因子红细胞 2 相关因子 1(也称为 Nfe2l1)和核呼吸因子(最初命名为-Pal)。因此,我们在这里确定 Nfe2l1 和-Pal 在核质呼吸和抗氧化转录谱的综合调节中发挥协同和拮抗作用。在小鼠胚胎成纤维细胞(MEFs)中,敲除导致-Pal 和 Nfe2l2 的表达水平显著降低,同时还包括 TFAM(线粒体转录因子 A)和其他靶基因。在 MEFs 中也观察到类似的抑制作用,但有一个例外,即 和 在 MEFs 中都明显上调。在 MEFs 中,Nfe2l1 和 Nfe2l2 对-Pal 和 TFAM 的正向调节的协同作用得到了验证。然而,人类-Pal 的表达不受 、 或 的影响,尽管 TFAM 被 Nfe2l1 激活但被 Nfe2l2 抑制;这种拮抗作用发生在 HepG2 细胞中。相反,几乎所有的小鼠 Nfe2l1、Nfe2l2 和共同靶基因在 MEFs 中都下调。相反,沉默-Pal 后,人 Nfe2l1、Nfe2l2 和相关报告基因上调,但外源表达-Pal 后下调。此外,Pitx2(pituitary homeobox 2)也被确定为 Nfe2l1 和 TFAM 的直接上游调节因子,除了-Pal。总的来说,这些 Nfe2l1、Nfe2l2 和-Pal 之间以及与 Pitx2 之间的交叉对话,从内质网到核质呼吸到线粒体通讯,都是为了靶向 TFAM,从而精细调节不同细胞氧化呼吸和抗氧化基因转录网络的稳健平衡,尽管它们在小鼠和人类之间存在差异。此外,值得注意的是,鉴于这些转录因子的相互调节,我们在解释从 Nfe2l1、Nfe2l2、-Pal 或 Pitx2 敲除或其功能获得突变体获得的相关实验结果时,应非常谨慎。