Department of Biosystems Science and Engineering, Eidgenössiche Technische Hochschule (ETH) Zürich, Mattenstr. 26, Basel, 4058, Switzerland.
Institute of Biophysics, Johannes Kepler University Linz, Gruberstr. 40, Linz, 4020, Austria.
BMC Biol. 2021 Jan 13;19(1):4. doi: 10.1186/s12915-020-00936-8.
Lipid-protein interactions stabilize protein oligomers, shape their structure, and modulate their function. Whereas in vitro experiments already account for the functional importance of lipids by using natural lipid extracts, in silico methods lack behind by embedding proteins in single component lipid bilayers. However, to accurately complement in vitro experiments with molecular details at very high spatio-temporal resolution, molecular dynamics simulations have to be performed in natural(-like) lipid environments.
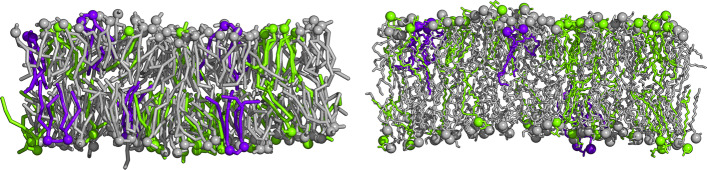
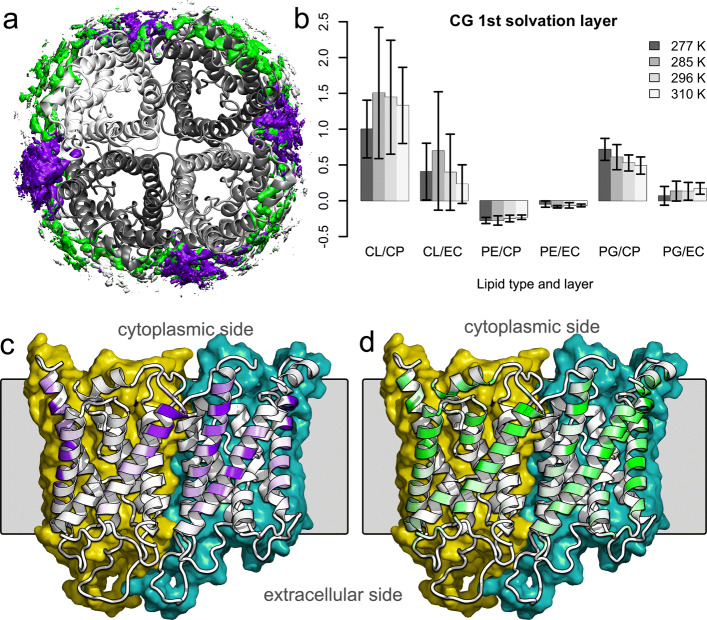
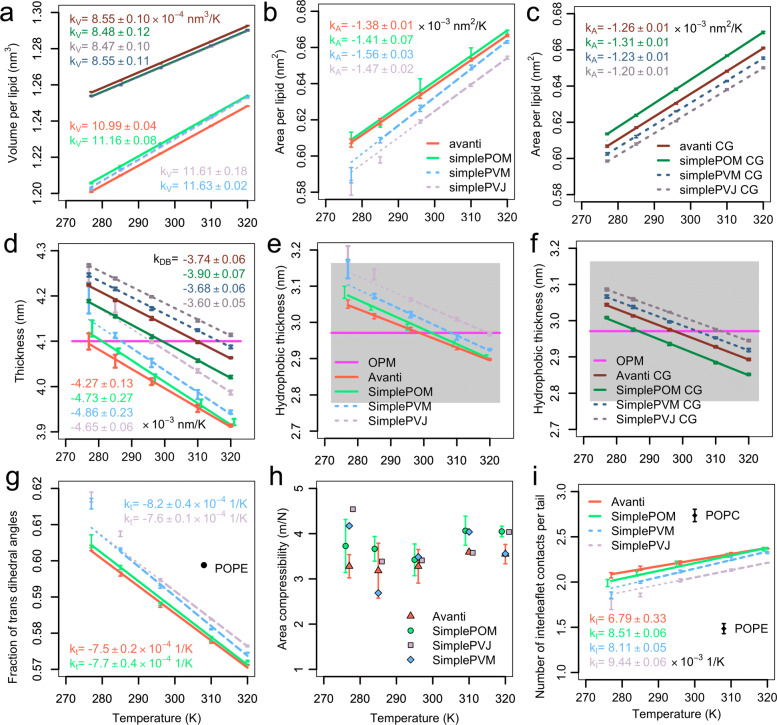
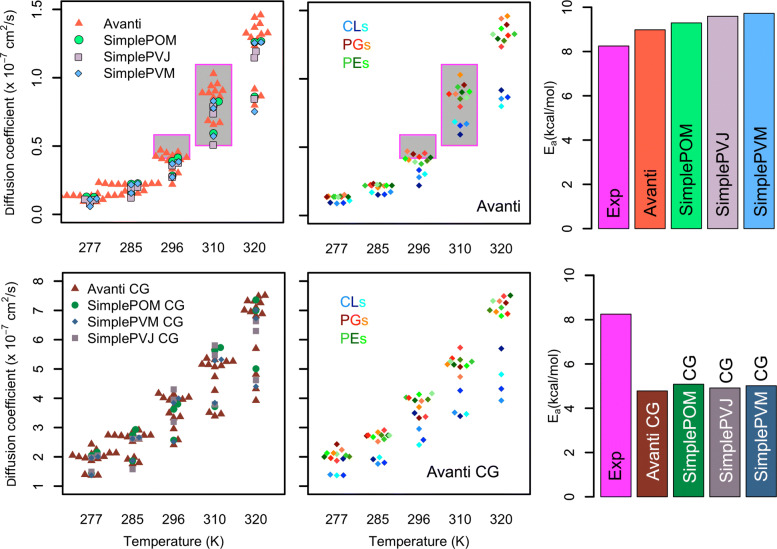
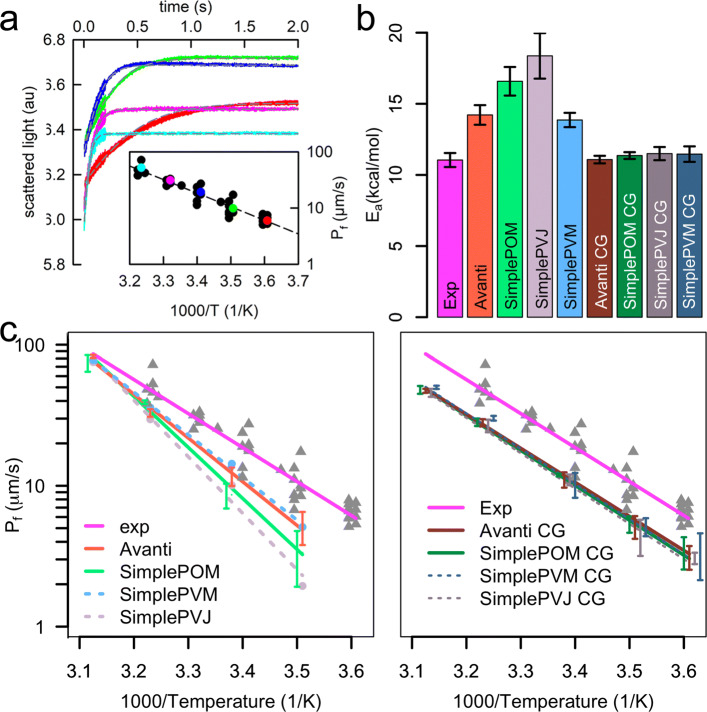
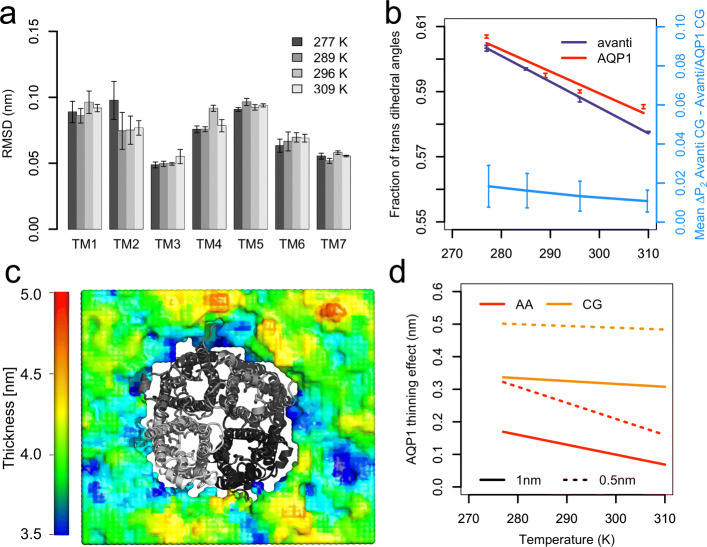
To enable more accurate MD simulations, we have prepared four membrane models of E. coli polar lipid extract, a typical model organism, each at all-atom (CHARMM36) and coarse-grained (Martini3) representations. These models contain all main lipid headgroup types of the E. coli inner membrane, i.e., phosphatidylethanolamines, phosphatidylglycerols, and cardiolipins, symmetrically distributed between the membrane leaflets. The lipid tail (un)saturation and propanylation stereochemistry represent the bacterial lipid tail composition of E. coli grown at 37C until 3/4 of the log growth phase. The comparison of the Simple three lipid component models to the complex 14-lipid component model Avanti over a broad range of physiologically relevant temperatures revealed that the balance of lipid tail unsaturation and propanylation in different positions and inclusion of lipid tails of various length maintain realistic values for lipid mobility, membrane area compressibility, lipid ordering, lipid volume and area, and the bilayer thickness. The only Simple model that was able to satisfactory reproduce most of the structural properties of the complex Avanti model showed worse agreement of the activation energy of basal water permeation with the here performed measurements. The Martini3 models reflect extremely well both experimental and atomistic behavior of the E. coli polar lipid extract membranes. Aquaporin-1 embedded in our native(-like) membranes causes partial lipid ordering and membrane thinning in its vicinity. Moreover, aquaporin-1 attracts and temporarily binds negatively charged lipids, mainly cardiolipins, with a distinct cardiolipin binding site in the crevice at the contact site between two monomers, most probably stabilizing the tetrameric protein assembly.
The here prepared and validated membrane models of E. coli polar lipids extract revealed that lipid tail complexity, in terms of double bond and cyclopropane location and varying lipid tail length, is key to stabilize membrane properties over a broad temperature range. In addition, they build a solid basis for manifold future simulation studies on more realistic lipid membranes bridging the gap between simulations and experiments.
脂质-蛋白质相互作用稳定蛋白质寡聚体,塑造其结构,并调节其功能。虽然体外实验已经通过使用天然脂质提取物来解释脂质的功能重要性,但由于在单组分脂质双层中嵌入蛋白质,因此计算方法落后了。然而,为了以非常高的时空分辨率用分子细节准确地补充体外实验,必须在天然(类似)脂质环境中进行分子动力学模拟。
为了能够进行更准确的 MD 模拟,我们制备了四种大肠杆菌极性脂质提取物的膜模型,这是一种典型的模型生物,每种模型都具有全原子(CHARMM36)和粗粒度(Martini3)表示。这些模型包含大肠杆菌内膜的所有主要脂质头基类型,即磷脂酰乙醇胺、磷脂酰甘油和心磷脂,对称分布在膜双层之间。脂质尾部(不)饱和度和丙酰化立体化学代表了在 37°C 下生长的大肠杆菌的细菌脂质尾部组成,直到对数生长阶段的 3/4。在广泛的生理相关温度范围内,将简单的三脂质成分模型与复杂的 14 脂质成分模型 Avanti 进行比较,结果表明,不同位置的脂质尾部不饱和度和丙酰化平衡以及各种长度的脂质尾部的包含可维持脂质流动性、膜面积压缩性、脂质有序性、脂质体积和面积以及双层厚度的现实值。唯一能够令人满意地再现复杂 Avanti 模型的大多数结构特性的简单模型显示,基底水渗透率的活化能与这里进行的测量结果的吻合度较差。Martini3 模型极好地反映了大肠杆菌极性脂质提取物膜的实验和原子行为。嵌入我们天然(类似)膜中的水通道蛋白-1会导致其附近的脂质部分有序化和膜变薄。此外,水通道蛋白-1吸引并暂时结合带负电荷的脂质,主要是心磷脂,在心磷脂结合位点在两个单体接触点之间的缝隙中,可能稳定四聚体蛋白组装。
这里制备和验证的大肠杆菌极性脂质提取物膜模型表明,脂质尾部的复杂性,包括双键和环丙烷的位置以及不同的脂质尾部长度,是在较宽温度范围内稳定膜性质的关键。此外,它们为在更现实的脂质膜上进行更多仿真研究奠定了坚实的基础,这些脂质膜缩小了仿真和实验之间的差距。