University of Murcia, IMIB-Arrixaca Institute of Biomedical Research, 30120, El Palmar, Murcia, Spain.
Department of Embryology, Carnegie Institution for Science, 3520 San Martin Drive, Baltimore, MD, 21218, USA.
Brain Struct Funct. 2021 Mar;226(2):519-562. doi: 10.1007/s00429-020-02197-1. Epub 2021 Jan 25.
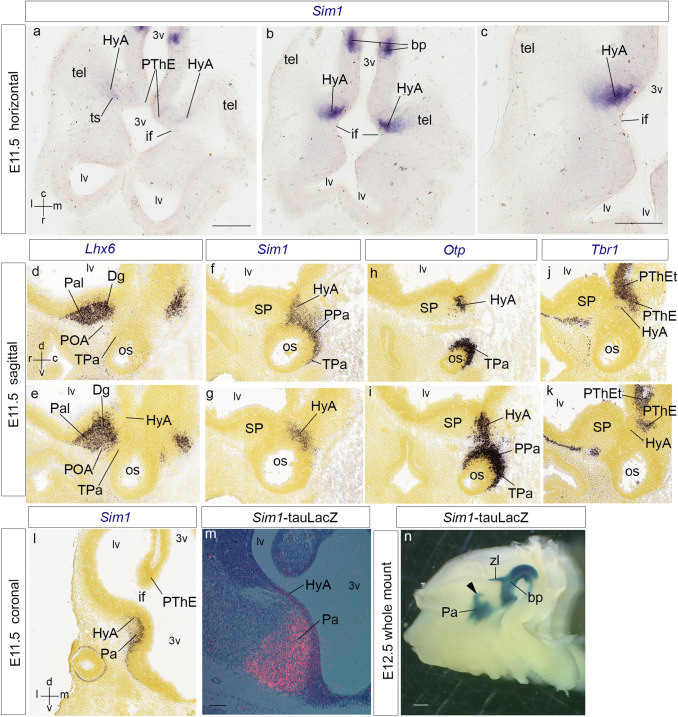
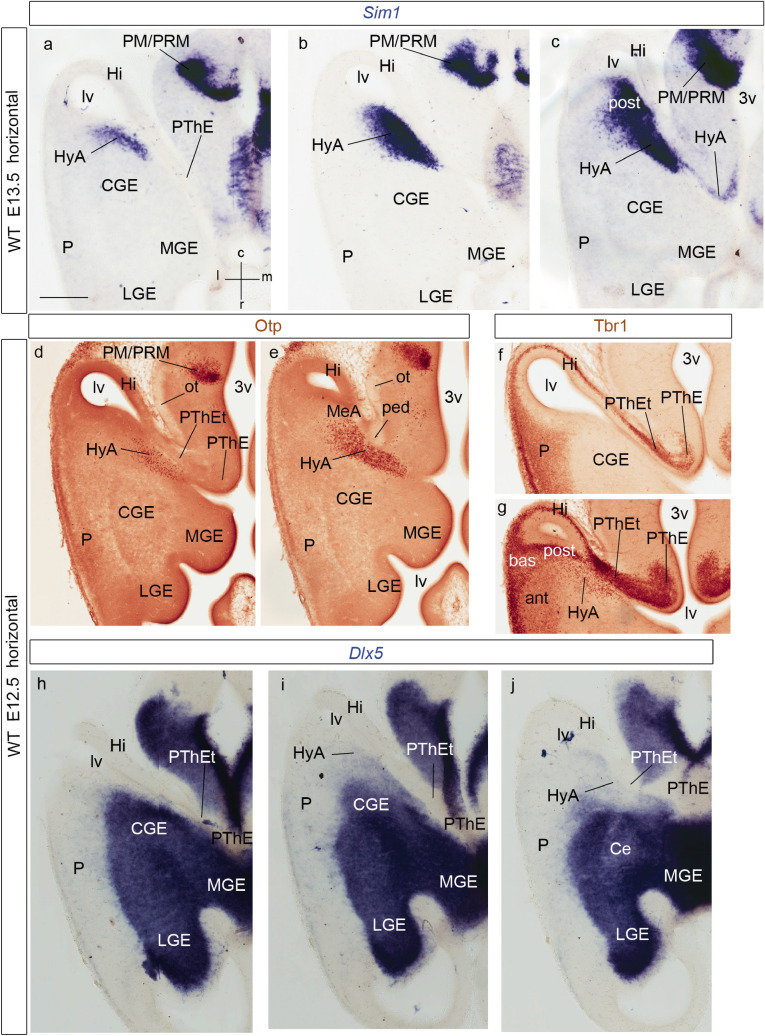
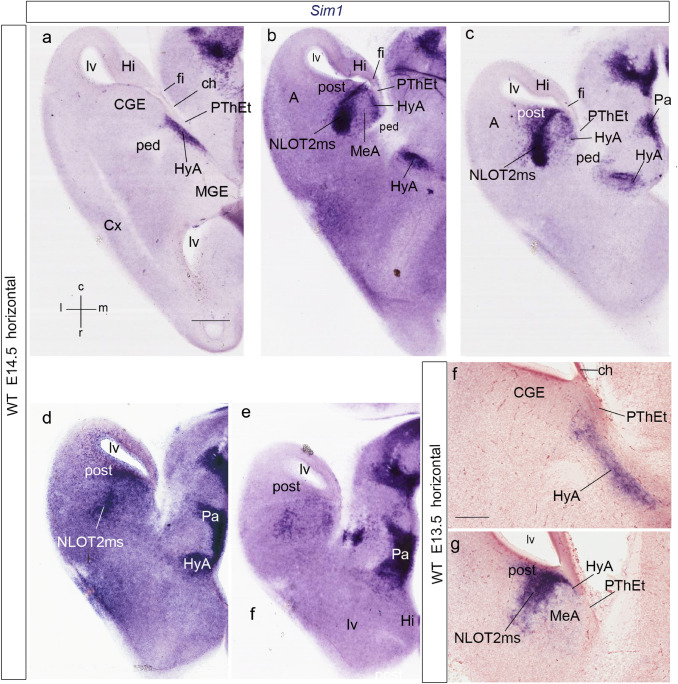
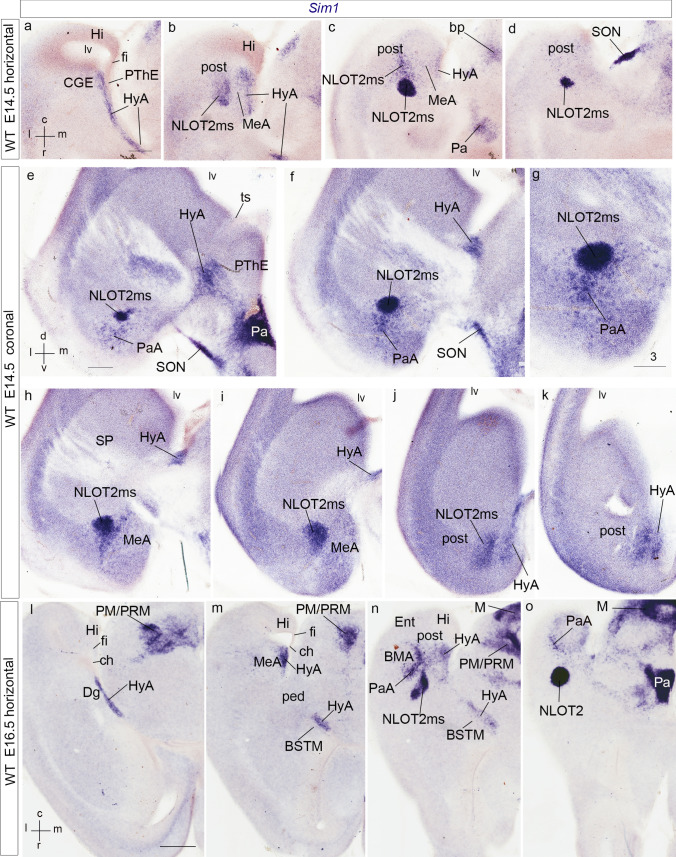
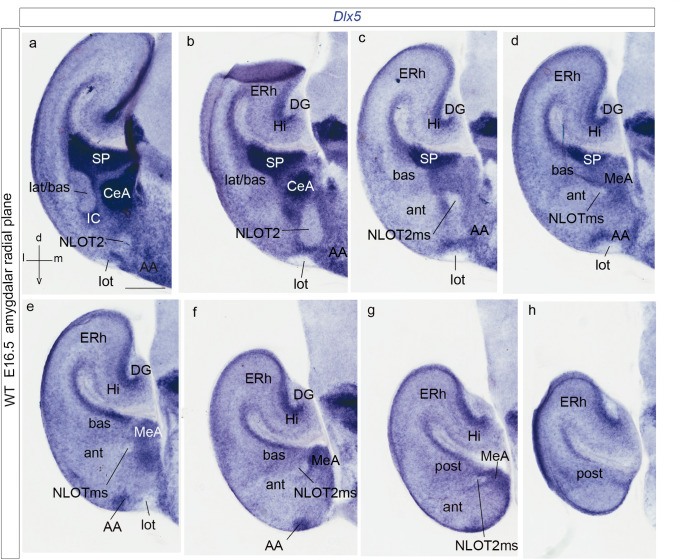
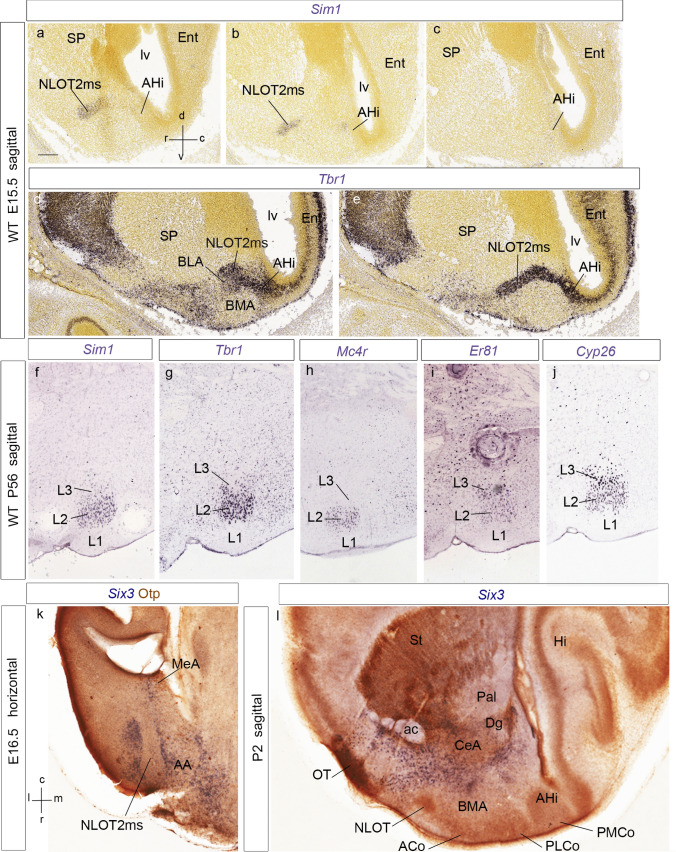
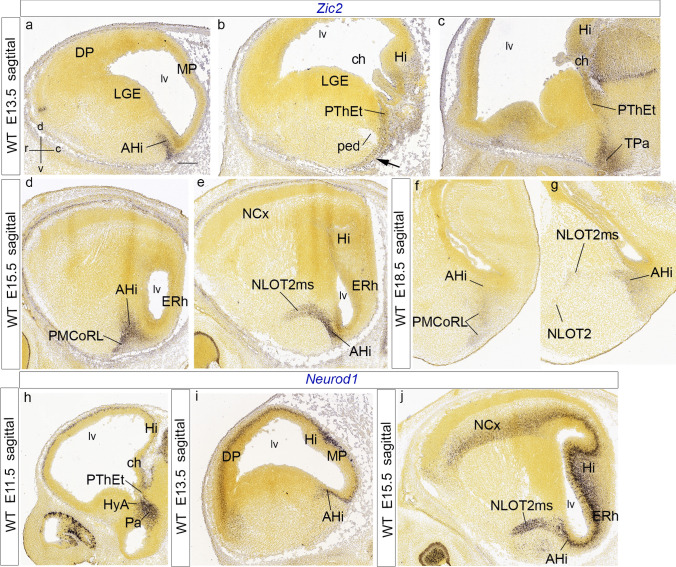
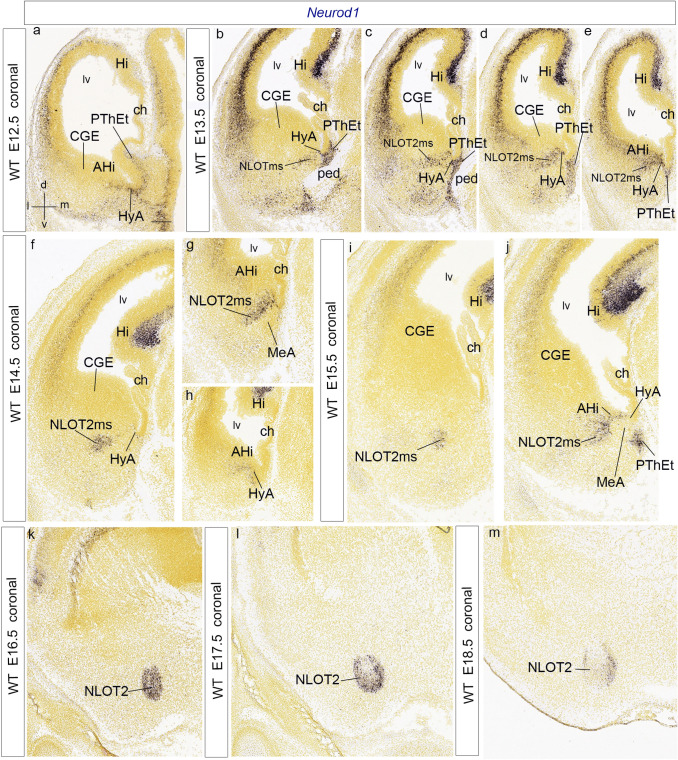
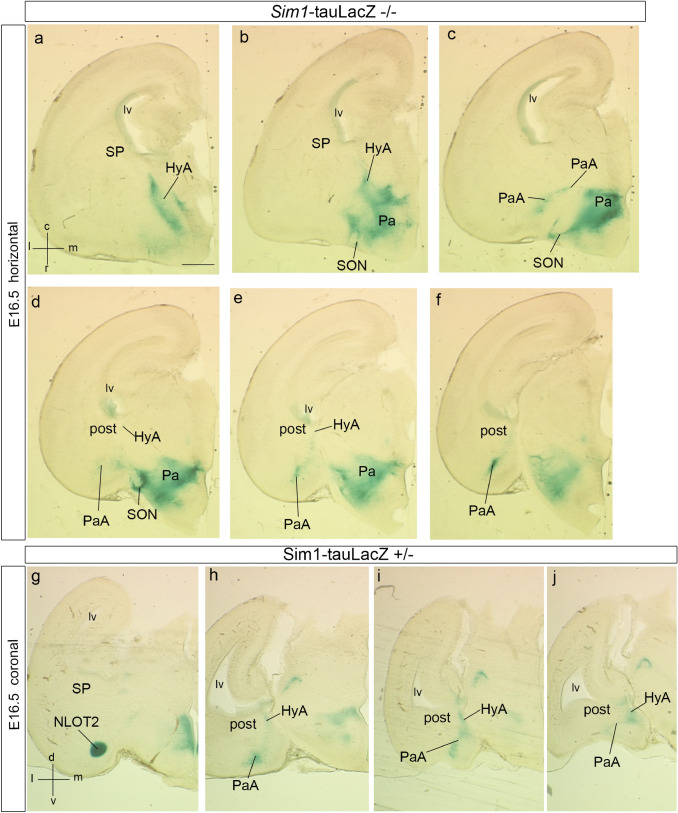
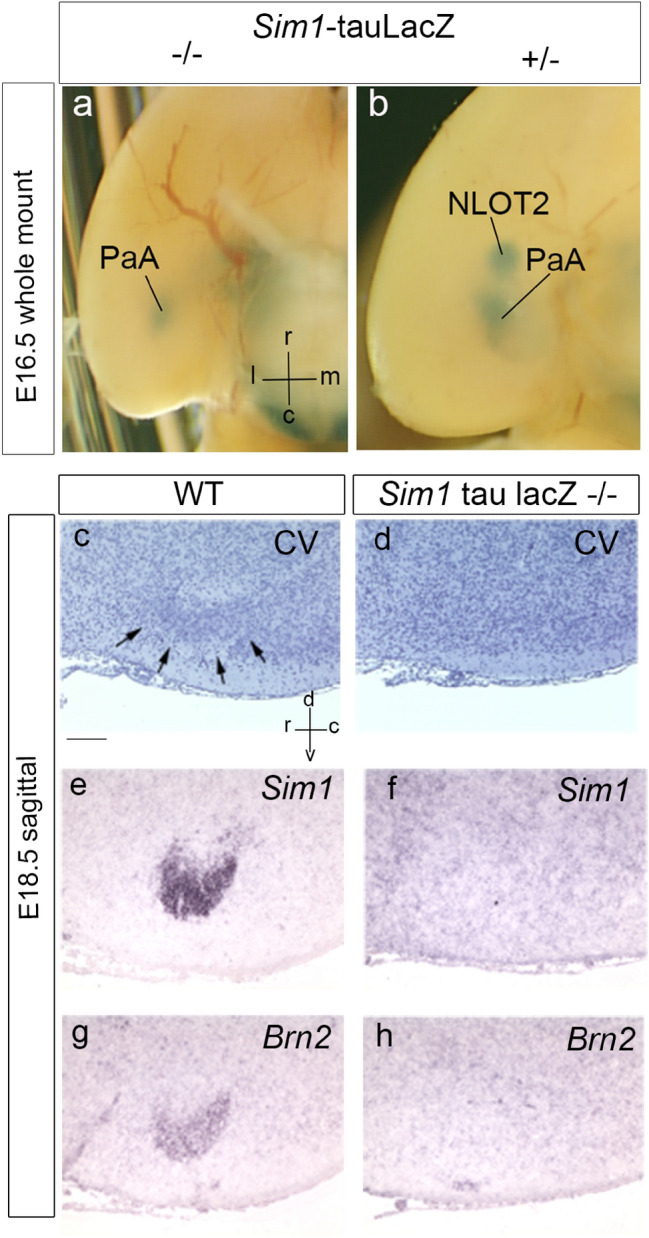
We focus this report on the nucleus of the lateral olfactory tract (NLOT), a superficial amygdalar nucleus receiving olfactory input. Mixed with its Tbr1-expressing layer 2 pyramidal cell population (NLOT2), there are Sim1-expressing cells whose embryonic origin and mode of arrival remain unclear. We examined this population with Sim1-ISH and a Sim1-tauLacZ mouse line. An alar hypothalamic origin is apparent at the paraventricular area, which expresses Sim1 precociously. This progenitor area shows at E10.5 a Sim1-expressing dorsal prolongation that crosses the telencephalic stalk and follows the terminal sulcus, reaching the caudomedial end of the pallial amygdala. We conceive this Sim1-expressing hypothalamo-amygdalar corridor (HyA) as an evaginated part of the hypothalamic paraventricular area, which participates in the production of Sim1-expressing cells. From E13.5 onwards, Sim1-expressing cells migrated via the HyA penetrate the posterior pallial amygdalar radial unit and associate therein to the incipient Tbr1-expressing migration stream which swings medially past the amygdalar anterior basolateral nucleus (E15.5), crosses the pallio-subpallial boundary (E16.5), and forms the NLOT2 within the anterior amygdala by E17.5. We conclude that the Tbr1-expressing NLOT2 cells arise strictly within the posterior pallial amygdalar unit, involving a variety of required gene functions we discuss. Our results are consistent with the experimental data on NLOT2 origin reported by Remedios et al. (Nat Neurosci 10:1141-1150, 2007), but we disagree on their implication in this process of the dorsal pallium, observed to be distant from the amygdala.
我们专注于本报告外侧嗅束(NLOT)的核心,这是一个接收嗅觉输入的浅层杏仁核。与表达 Tbr1 的第 2 层锥体神经元混合,还有表达 Sim1 的细胞,但其胚胎起源和到达方式尚不清楚。我们使用 Sim1-ISH 和 Sim1-tauLacZ 小鼠系检查了这一群细胞。在室旁区可见明显的翼状下丘脑起源,该区域过早表达 Sim1。这个前体细胞区在 E10.5 时显示出表达 Sim1 的背侧延伸,穿过端脑柄并沿着终沟延伸,到达皮质杏仁核的尾内侧端。我们将这个表达 Sim1 的下丘脑-杏仁核走廊(HyA)视为下丘脑室旁区的一个外生部分,它参与了表达 Sim1 的细胞的产生。从 E13.5 开始,通过 HyA 迁移的表达 Sim1 的细胞穿透后皮质杏仁核放射状单位,并在其中与初始表达 Tbr1 的迁移流相关联,该迁移流在杏仁核前基底外侧核(E15.5)处向内侧摆动,穿过皮质-下皮质边界(E16.5),并在 E17.5 时在前杏仁核内形成 NLOT2。我们得出结论,表达 Tbr1 的 NLOT2 细胞严格起源于后皮质杏仁核单元,涉及我们讨论的各种必需基因功能。我们的结果与 Remedios 等人报道的 NLOT2 起源的实验数据一致(Nat Neurosci 10:1141-1150, 2007),但我们不同意他们关于这个过程的背侧皮质在这个过程中的作用的观点,观察到它远离杏仁核。