Institute of Molecular Biology, Department of Biology, University of Oregon, 1229 Franklin Boulevard, Eugene, OR 97403, USA.
Department of Epigenetics and Molecular Carcinogenesis, MD Anderson Cancer Center, 1808 Park Road 1C, Smithville, TX 78957, USA.
Curr Biol. 2021 Apr 12;31(7):1508-1514.e5. doi: 10.1016/j.cub.2021.03.008. Epub 2021 Mar 18.
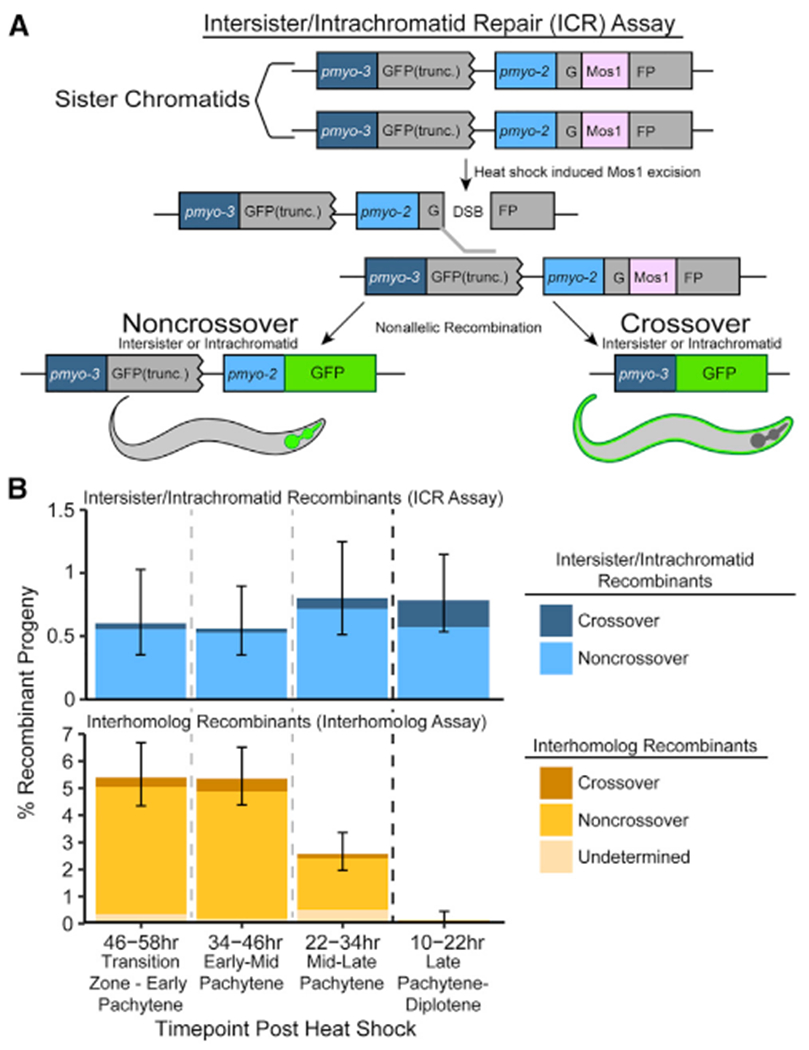
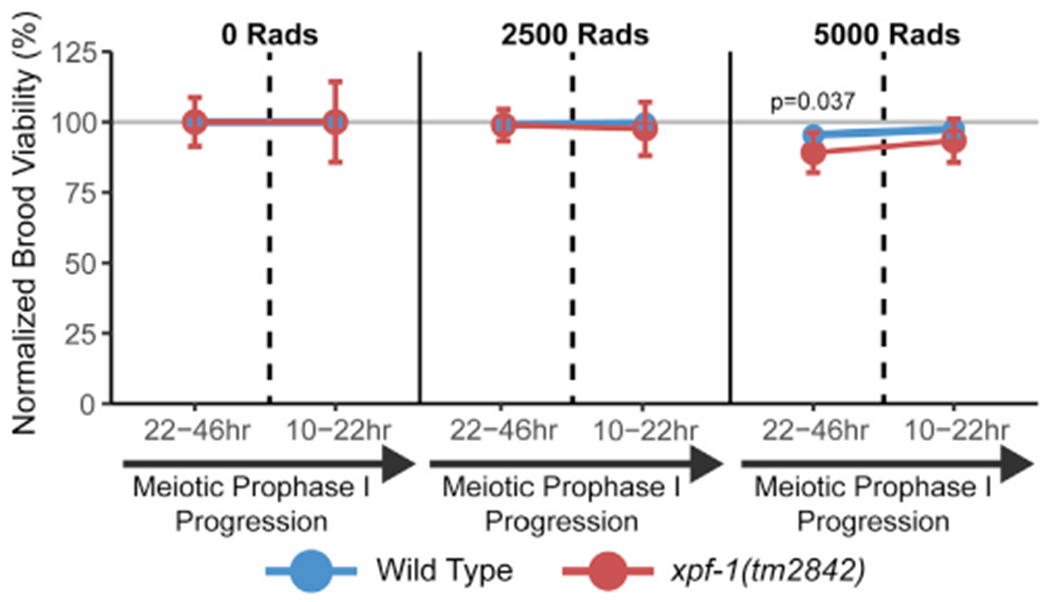
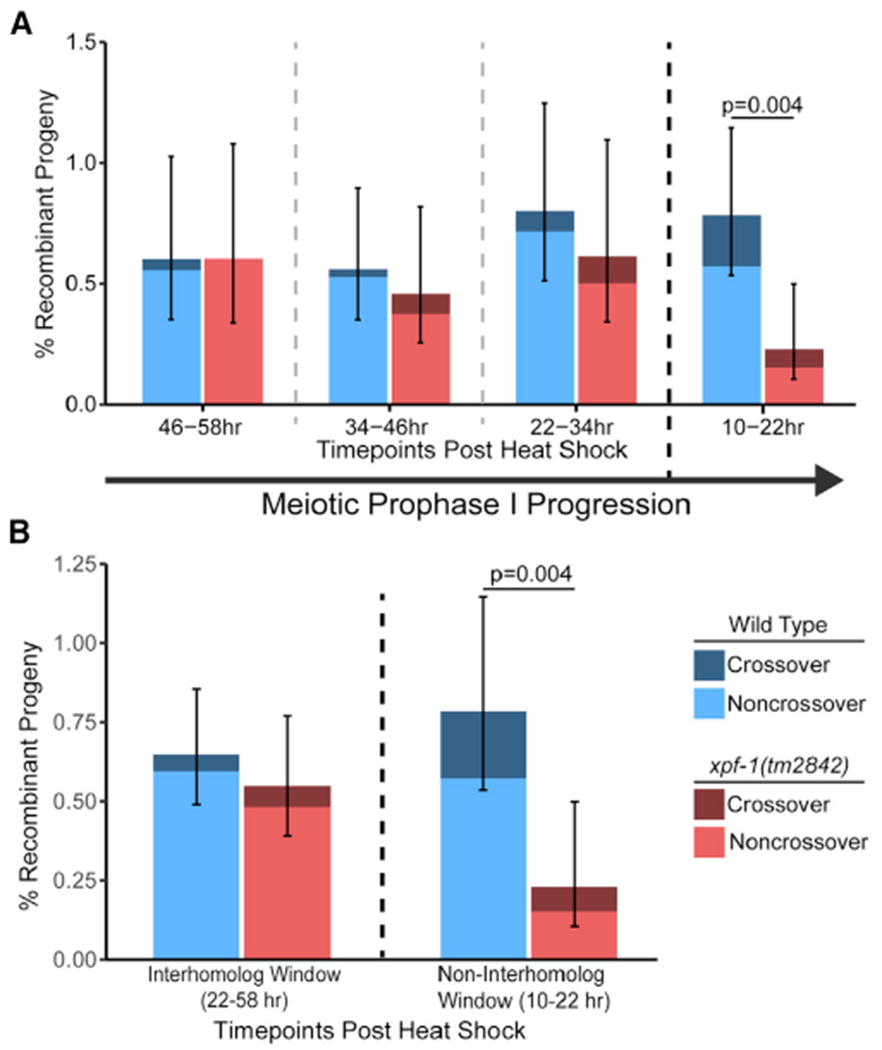
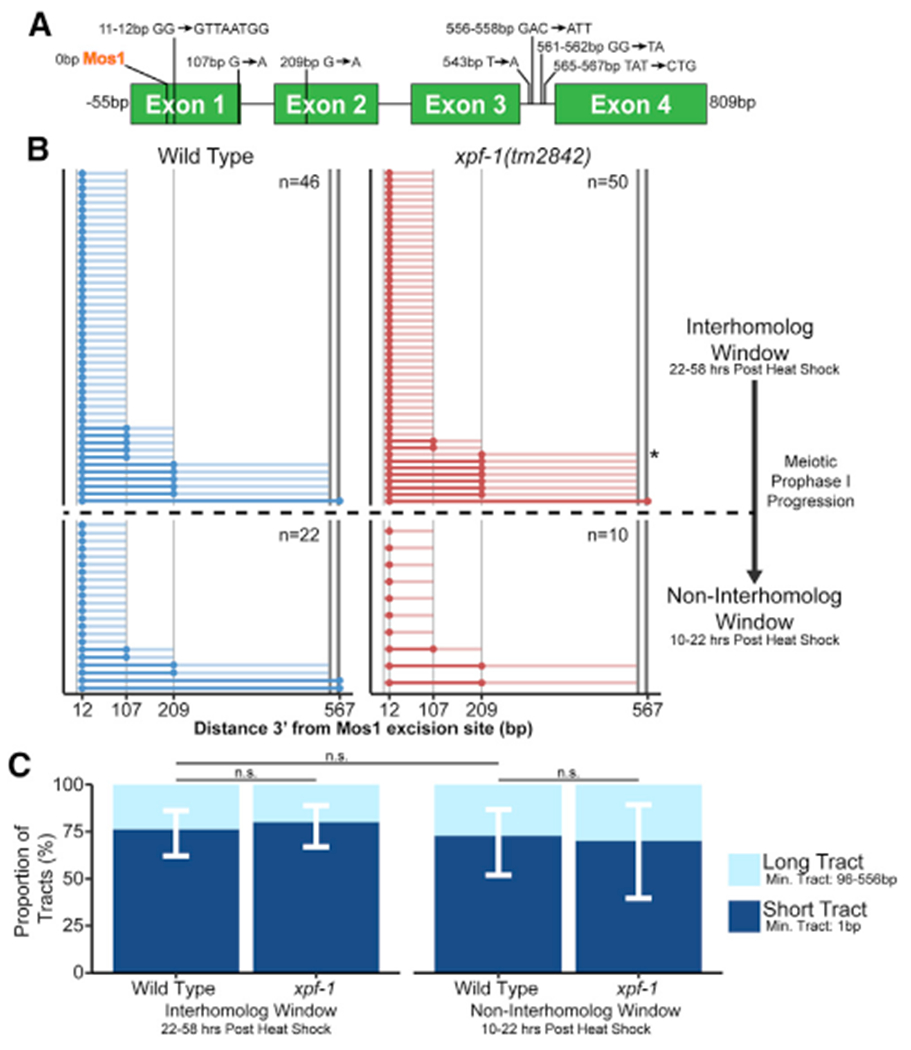
During meiosis, the maintenance of genome integrity is critical for generating viable haploid gametes. In meiotic prophase I, double-strand DNA breaks (DSBs) are induced and a subset of these DSBs are repaired as interhomolog crossovers to ensure proper chromosome segregation. DSBs not resolved as crossovers with the homolog must be repaired by other pathways to ensure genome integrity. To determine if alternative repair templates can be engaged for meiotic DSB repair during oogenesis, we developed an assay to detect sister and/or intra-chromatid repair events at a defined DSB site during Caenorhabditis elegans meiosis. Using this assay, we directly demonstrate that the sister chromatid or the same DNA molecule can be engaged as a meiotic repair template for both crossover and noncrossover recombination, with noncrossover events being the predominant recombination outcome. We additionally find that the sister or intra-chromatid substrate is available as a recombination partner for DSBs induced throughout meiotic prophase I, including late prophase when the homolog is unavailable. Analysis of noncrossover conversion tract sequences reveals that DSBs are processed similarly throughout prophase I. We further present data indicating that the XPF-1 nuclease functions in late prophase to promote sister or intra-chromatid repair at steps of recombination following joint molecule processing. Despite its function in sister or intra-chromatid repair, we find that xpf-1 mutants do not exhibit severe defects in progeny viability following exposure to ionizing radiation. Overall, we propose that C. elegans XPF-1 may assist as an intersister or intrachromatid resolvase only in late prophase I.
在减数分裂过程中,维持基因组完整性对于产生可行的单倍体配子至关重要。在减数分裂前期 I 中,会诱导双链 DNA 断裂 (DSB),其中一部分 DSB 作为同源染色体交叉被修复,以确保染色体正确分离。未被同源染色体交叉修复的 DSB 必须通过其他途径修复,以确保基因组完整性。为了确定在卵子发生过程中是否可以利用替代修复模板来修复减数分裂 DSB,我们开发了一种测定方法,用于在秀丽隐杆线虫减数分裂期间检测特定 DSB 位点的姐妹染色单体和/或染色内修复事件。使用该测定方法,我们直接证明姐妹染色单体或同一 DNA 分子可作为同源染色体交叉和非交叉重组的减数修复模板,而非交叉事件是主要的重组结果。我们还发现,当同源染色体不可用时,姐妹或染色内底物可作为减数分裂前期 I 中诱导的 DSB 的重组伙伴,包括前期后期。对非交叉转换区序列的分析表明,DSB 在整个前期 I 中以相似的方式进行处理。我们进一步提供的数据表明,XPF-1 核酸内切酶在前期后期发挥作用,以促进联合分子处理后重组步骤中的姐妹或染色内修复。尽管 XPF-1 在姐妹或染色内修复中发挥作用,但我们发现 XPF-1 突变体在暴露于电离辐射后不会导致后代活力严重缺陷。总的来说,我们提出 C. elegans XPF-1 可能仅在前期后期作为姐妹或染色内解旋酶来辅助。