University College London, Department of Chemistry, London, United Kingdom.
University College London School of Pharmacy, Pharmaceutical and Biological Chemistry, London, United Kingdom.
Elife. 2021 Mar 23;10:e66567. doi: 10.7554/eLife.66567.
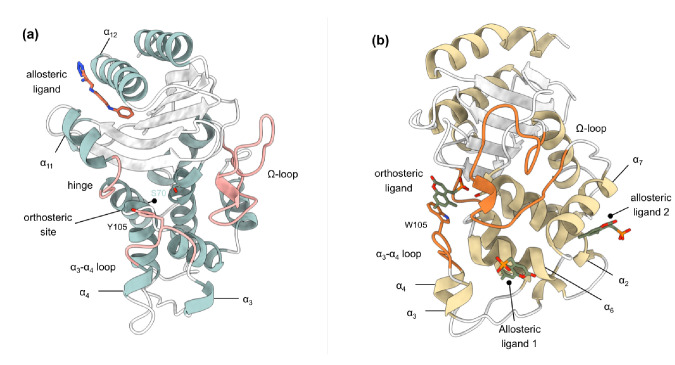
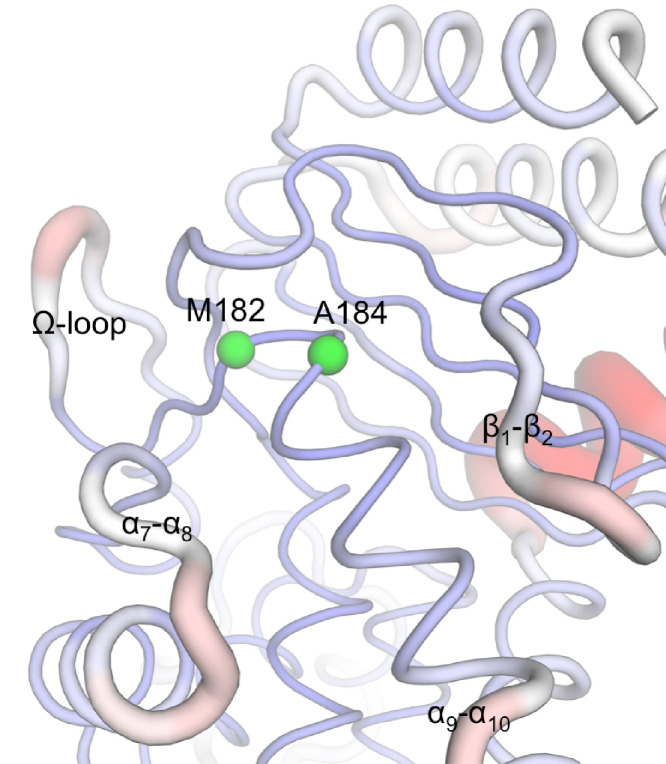
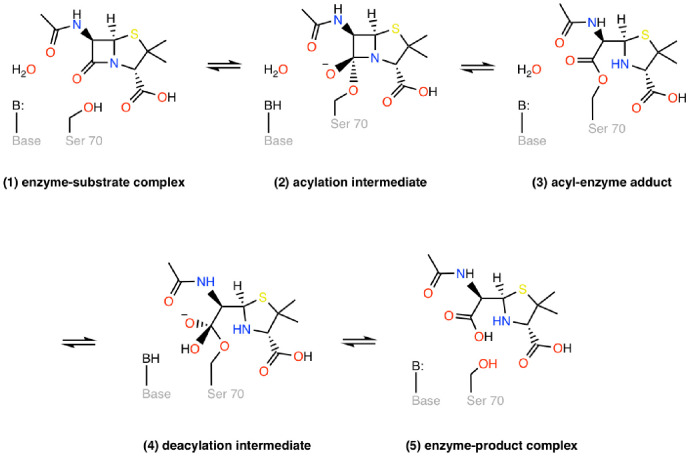
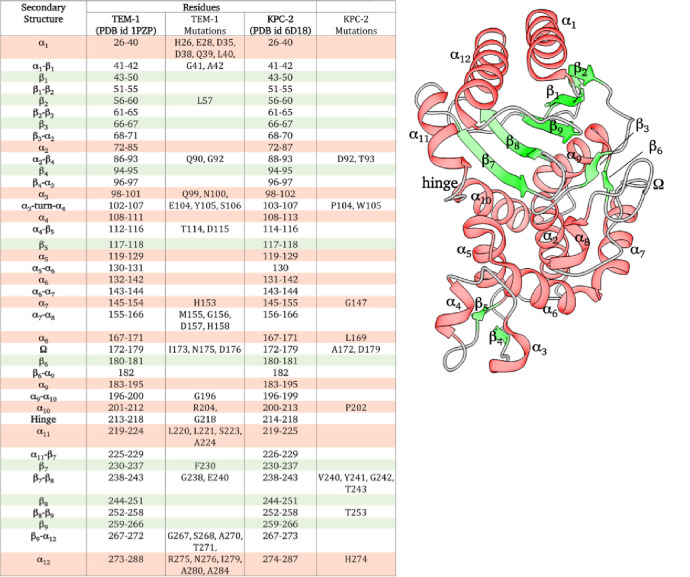
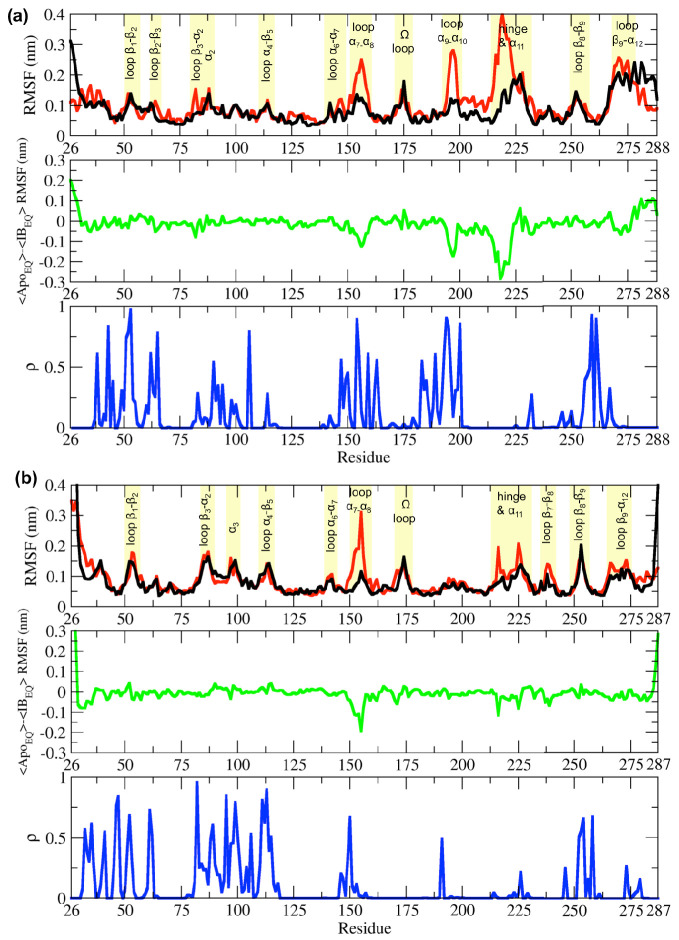
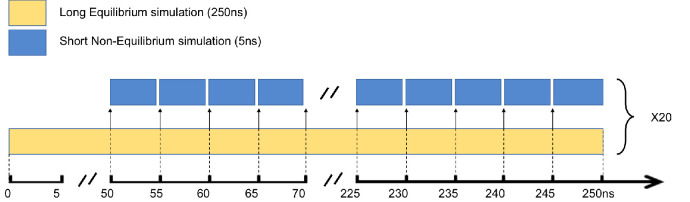
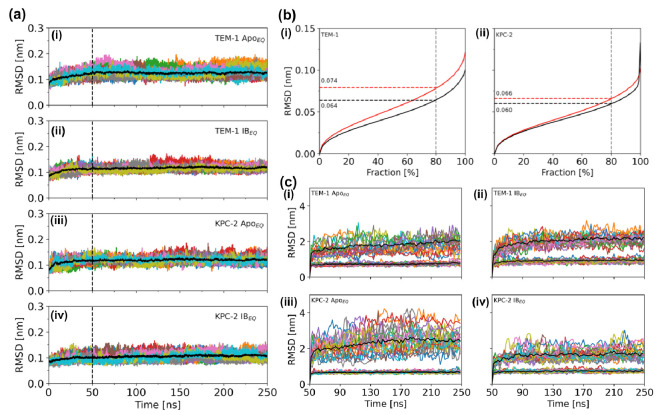
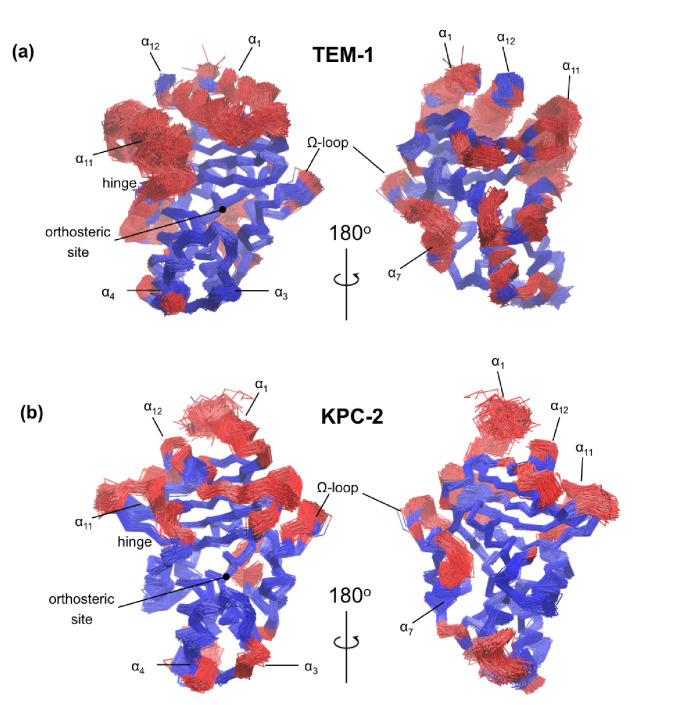
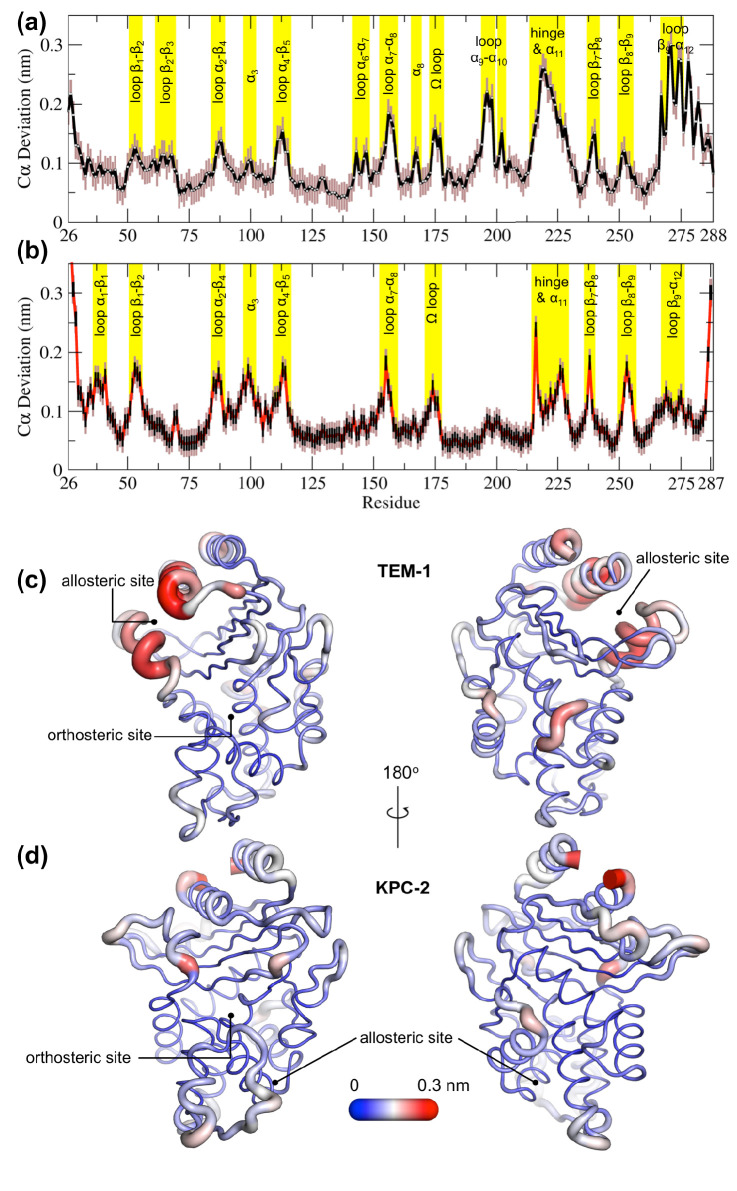
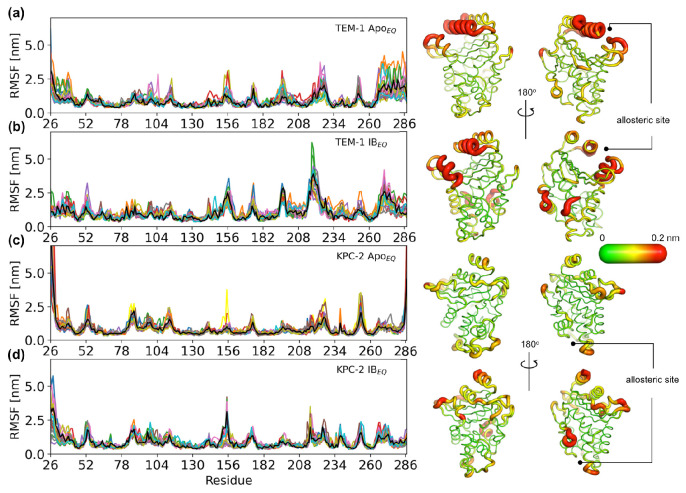
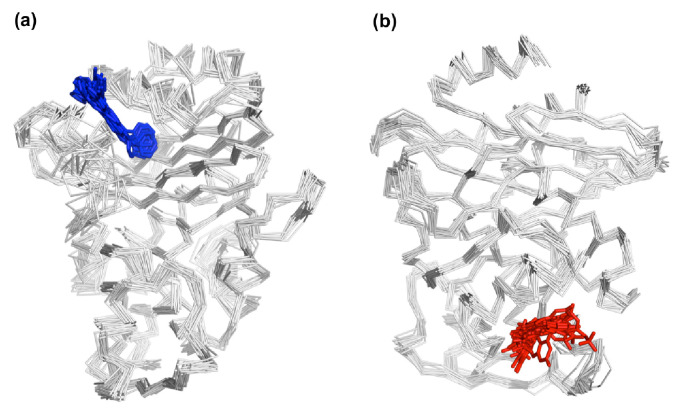
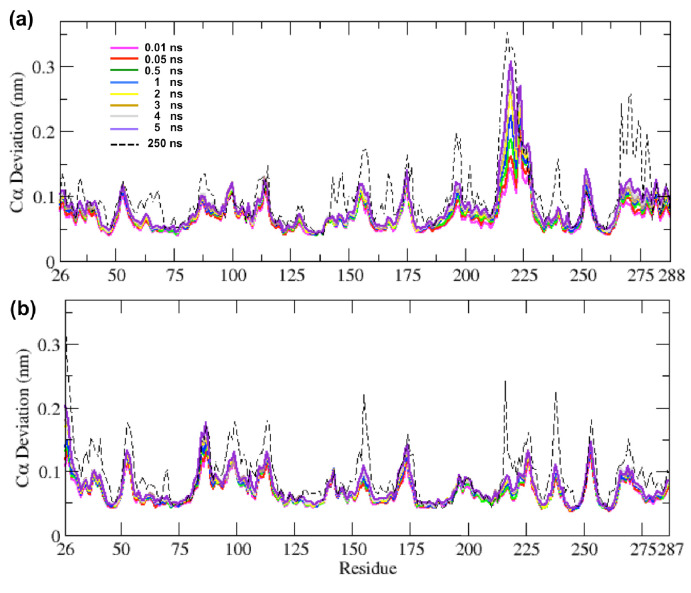
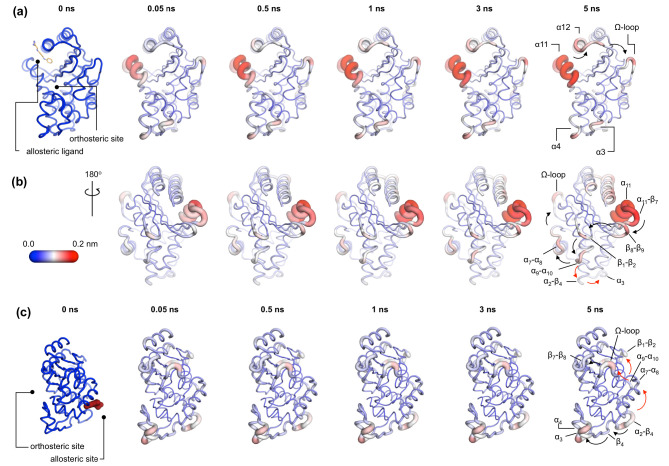
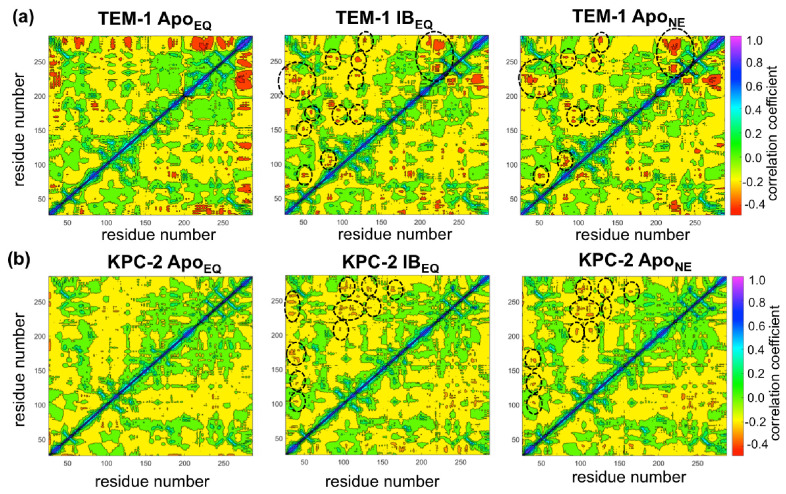
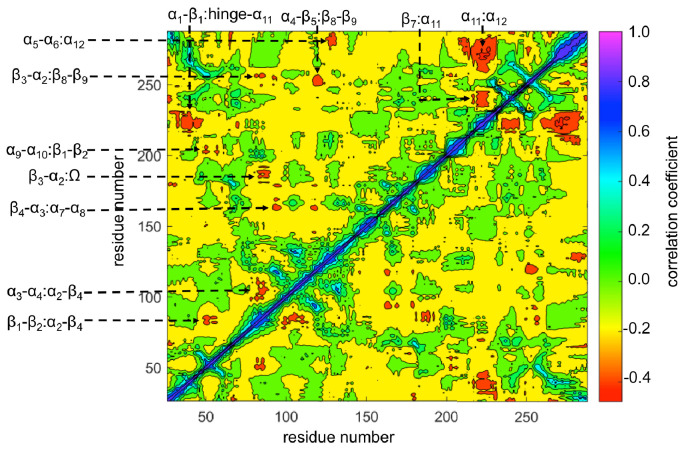
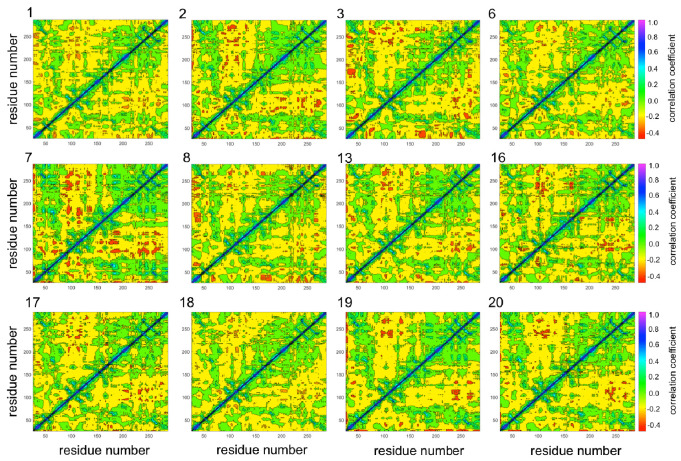
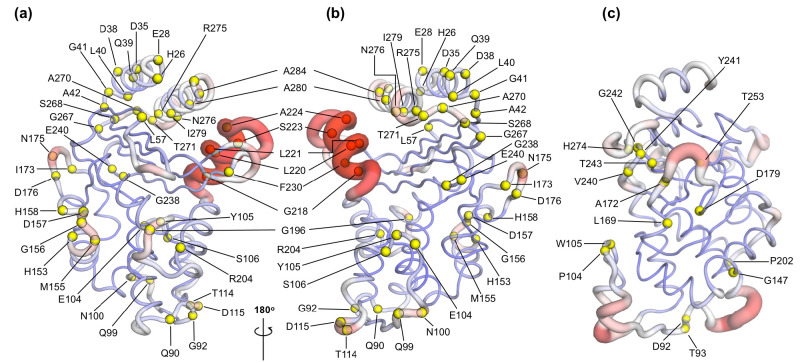
Understanding allostery in enzymes and tools to identify it offer promising alternative strategies to inhibitor development. Through a combination of equilibrium and nonequilibrium molecular dynamics simulations, we identify allosteric effects and communication pathways in two prototypical class A β-lactamases, TEM-1 and KPC-2, which are important determinants of antibiotic resistance. The nonequilibrium simulations reveal pathways of communication operating over distances of 30 Å or more. Propagation of the signal occurs through cooperative coupling of loop dynamics. Notably, 50% or more of clinically relevant amino acid substitutions map onto the identified signal transduction pathways. This suggests that clinically important variation may affect, or be driven by, differences in allosteric behavior, providing a mechanism by which amino acid substitutions may affect the relationship between spectrum of activity, catalytic turnover, and potential allosteric behavior in this clinically important enzyme family. Simulations of the type presented here will help in identifying and analyzing such differences.
理解酶的变构作用以及识别它的工具为抑制剂开发提供了有前景的替代策略。通过平衡和非平衡分子动力学模拟的组合,我们在两种典型的 A 类β-内酰胺酶(TEM-1 和 KPC-2)中鉴定出变构效应和通讯途径,这些是抗生素耐药性的重要决定因素。非平衡模拟揭示了在 30Å 或更长距离上起作用的通讯途径。信号的传播是通过环动力学的协同耦合发生的。值得注意的是,50%或更多的临床相关氨基酸取代映射到鉴定的信号转导途径上。这表明临床重要的变异可能会影响或由变构行为的差异驱动,为氨基酸取代如何影响该临床重要酶家族的活性谱、催化周转率和潜在变构行为之间的关系提供了一种机制。这里提出的模拟将有助于识别和分析此类差异。