School of Life Science and Technology, ShanghaiTech University, Shanghai, China.
College of Life and Environmental Sciences, Hangzhou Normal University, Hangzhou, China.
Elife. 2021 May 5;10:e67628. doi: 10.7554/eLife.67628.
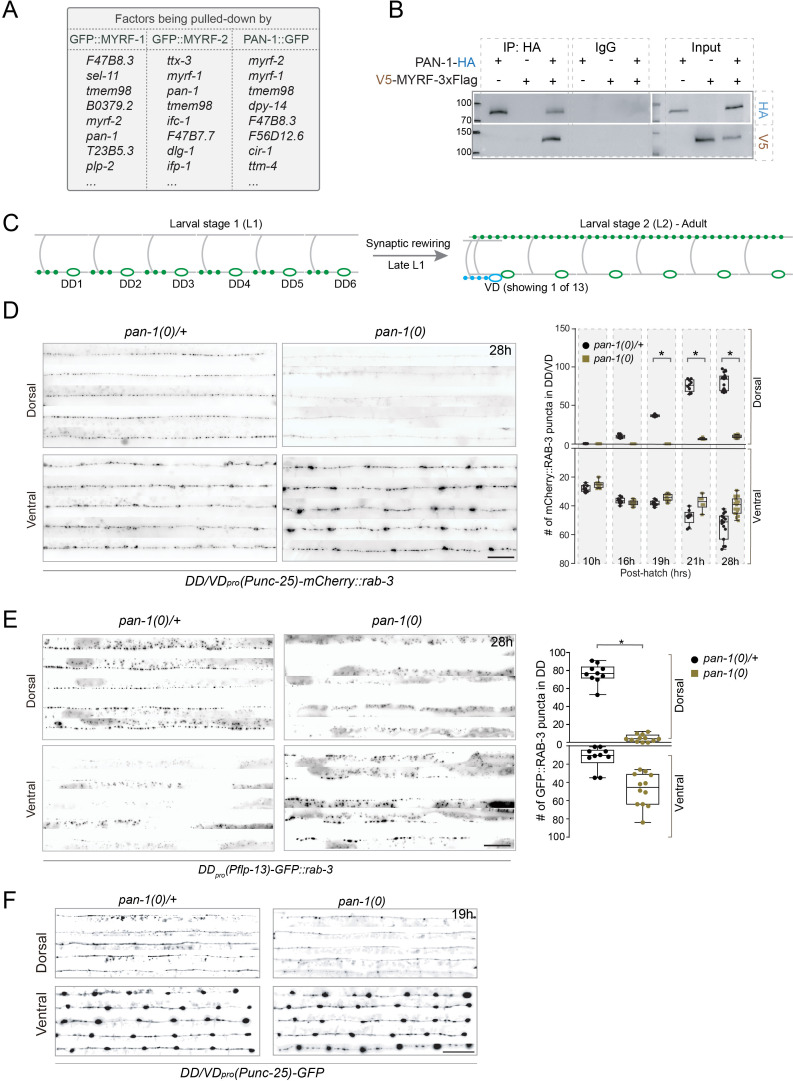
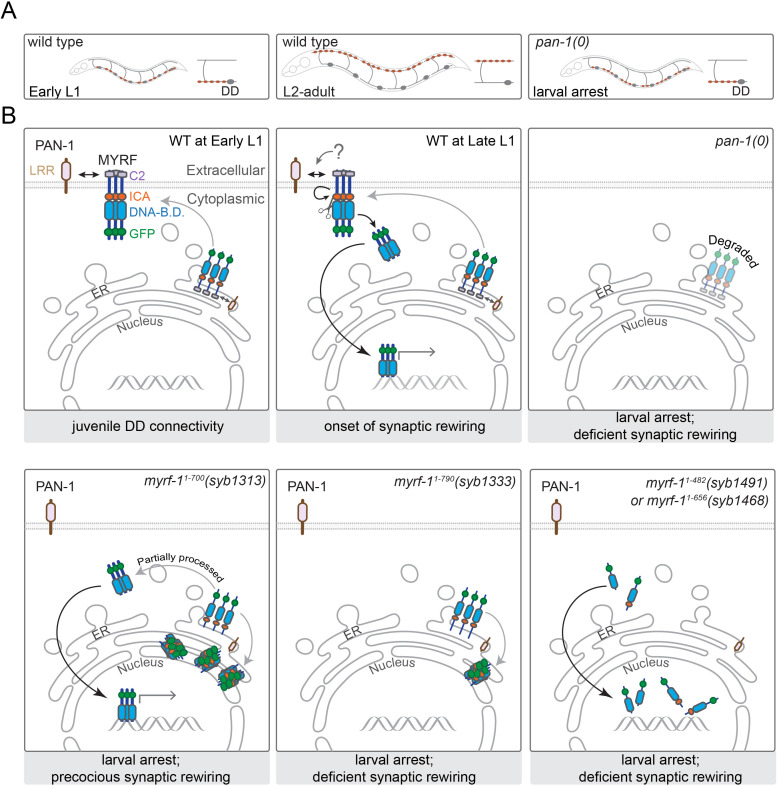
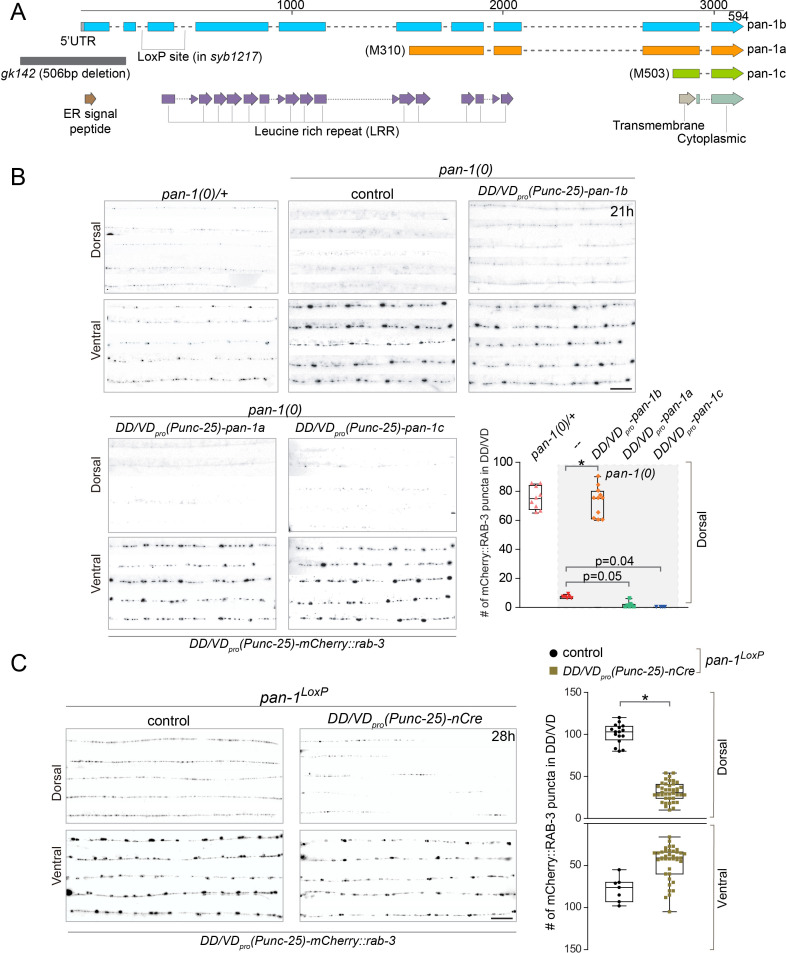
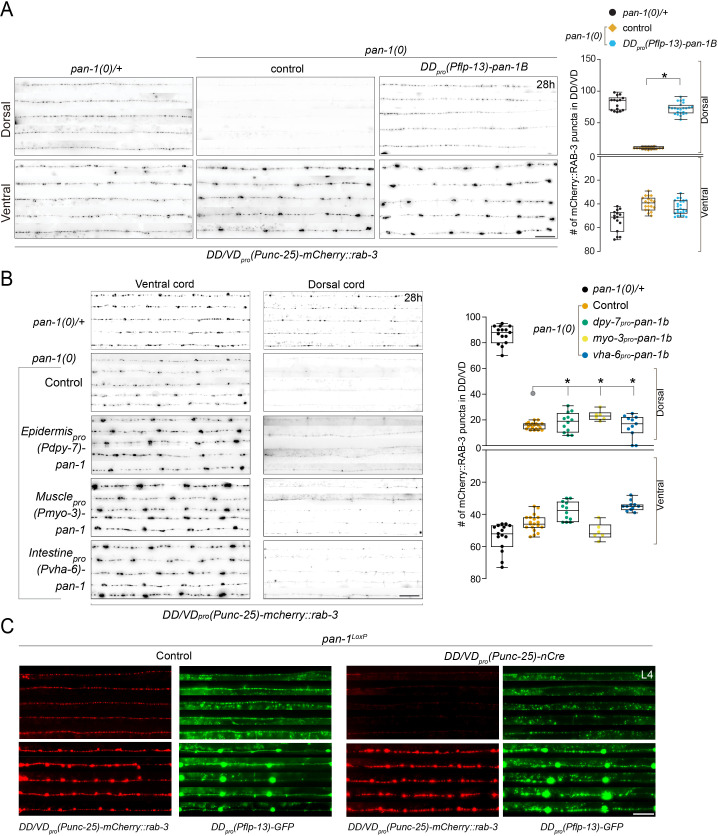
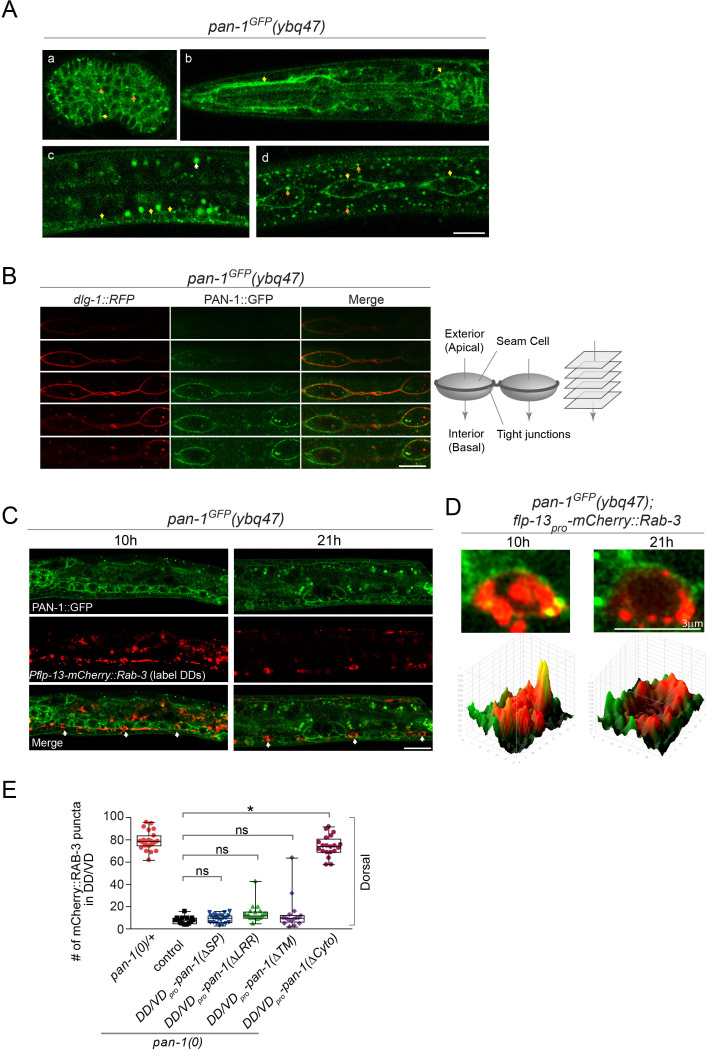
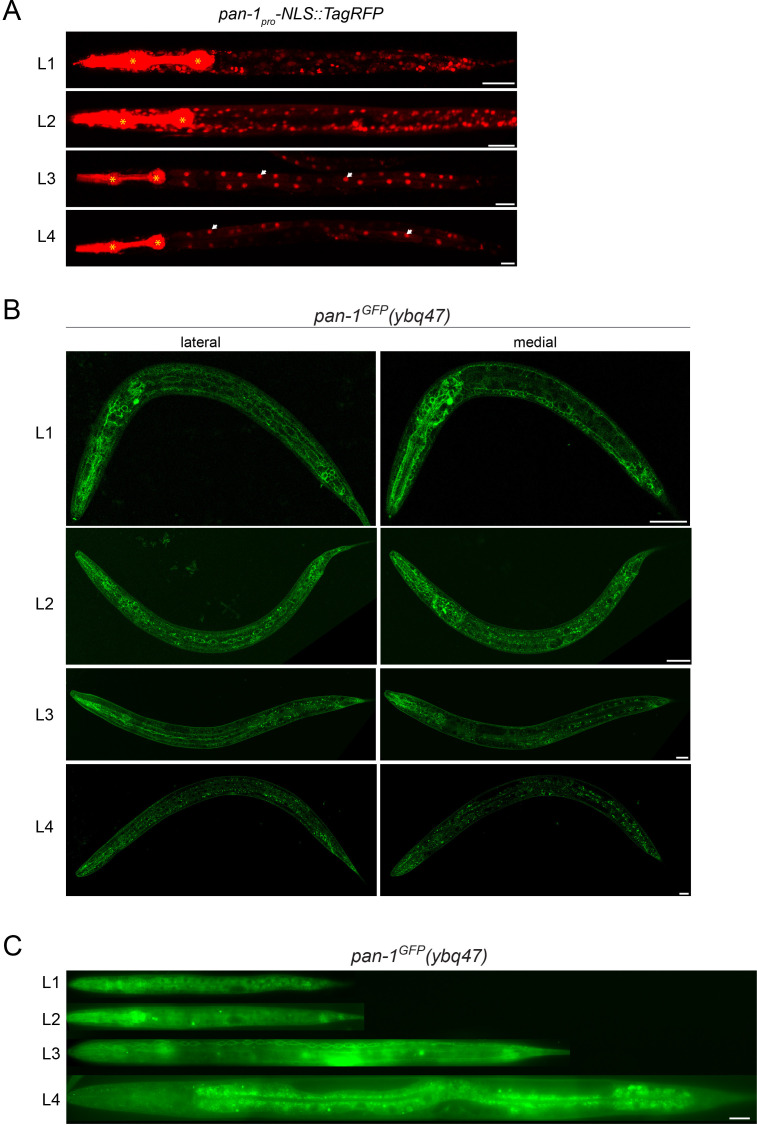
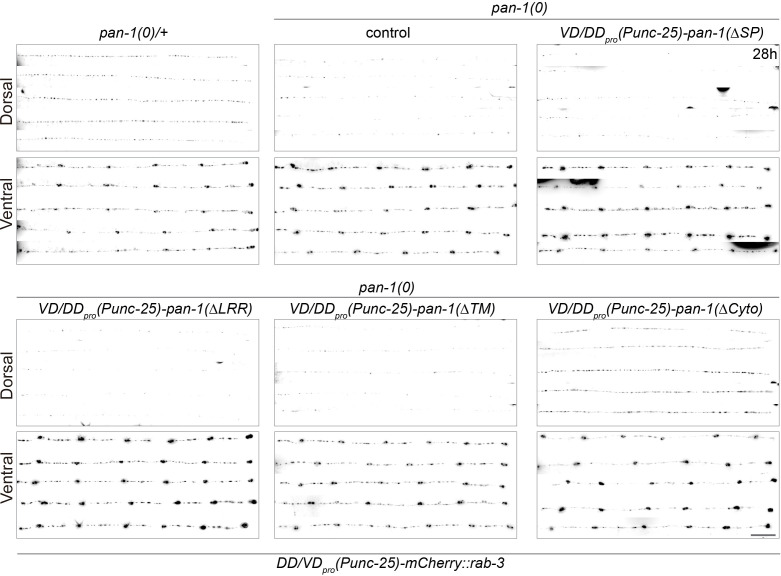
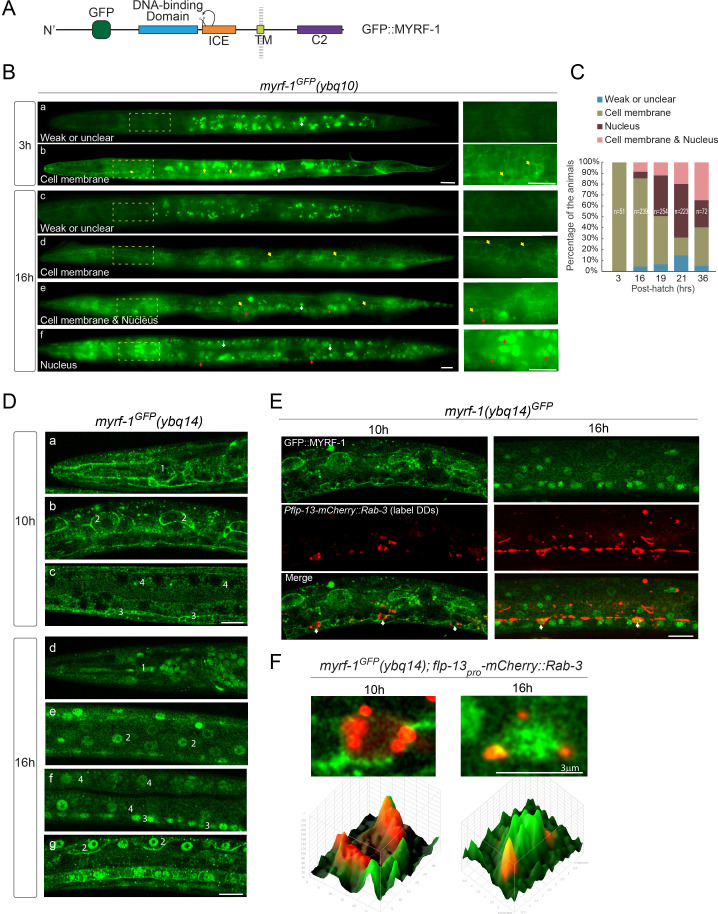
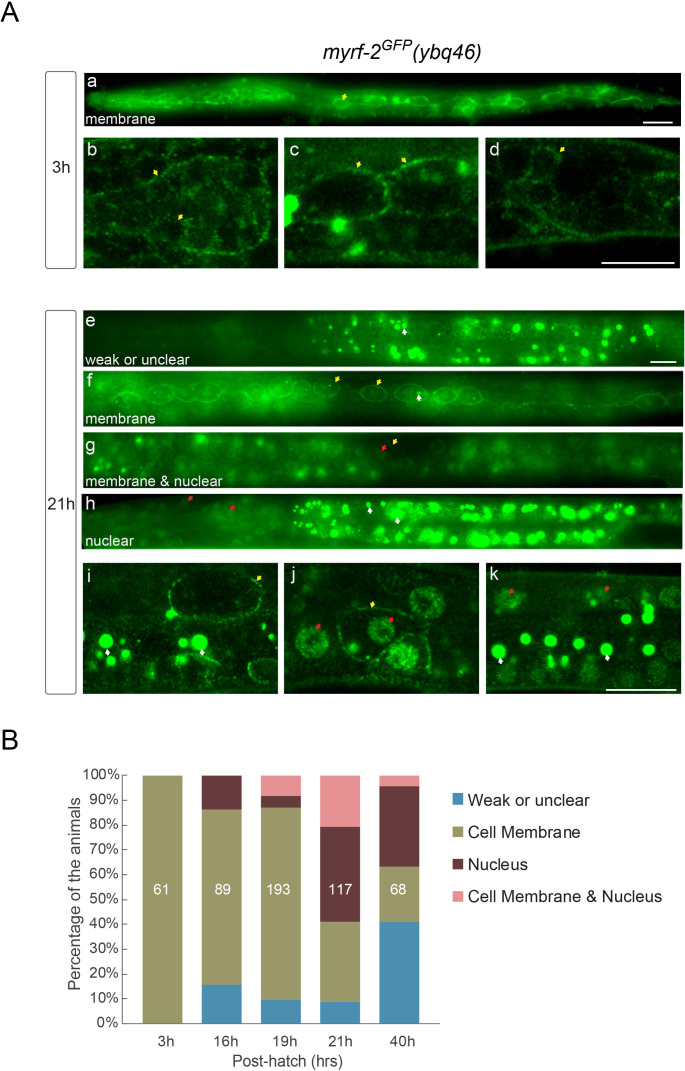
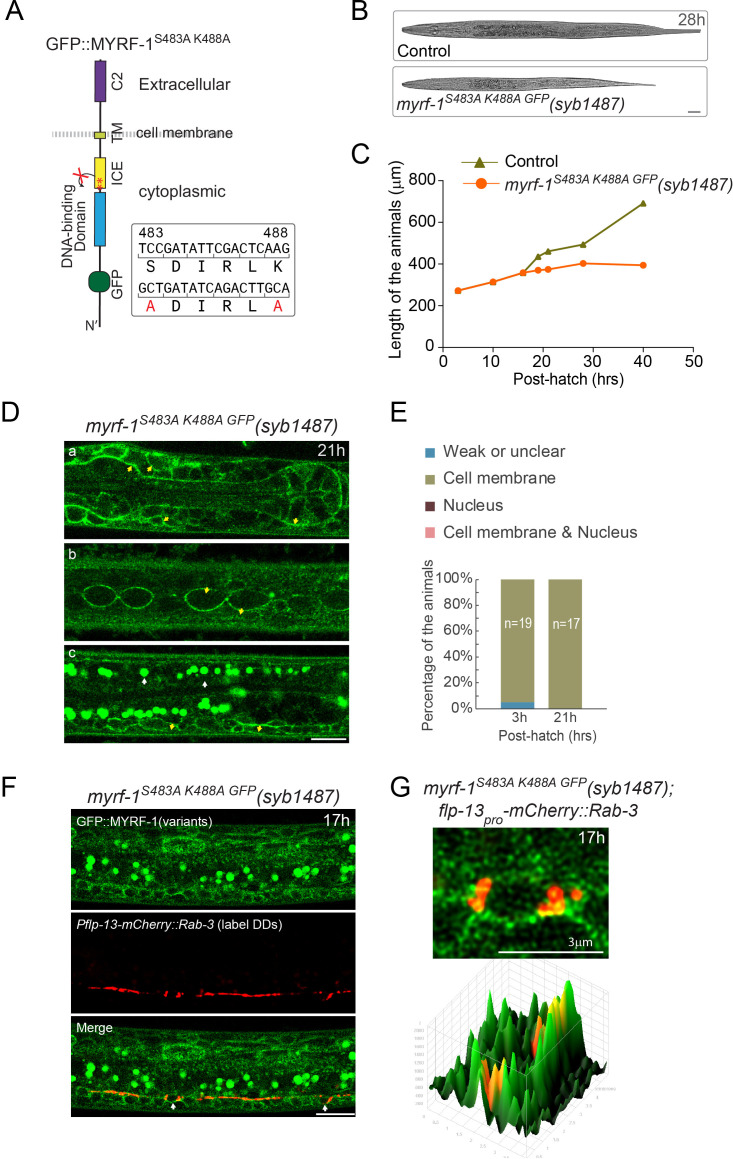
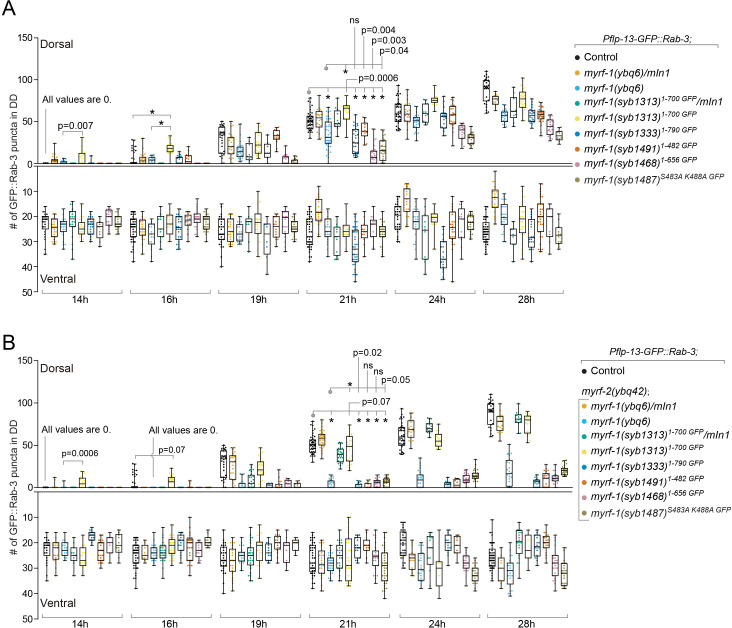
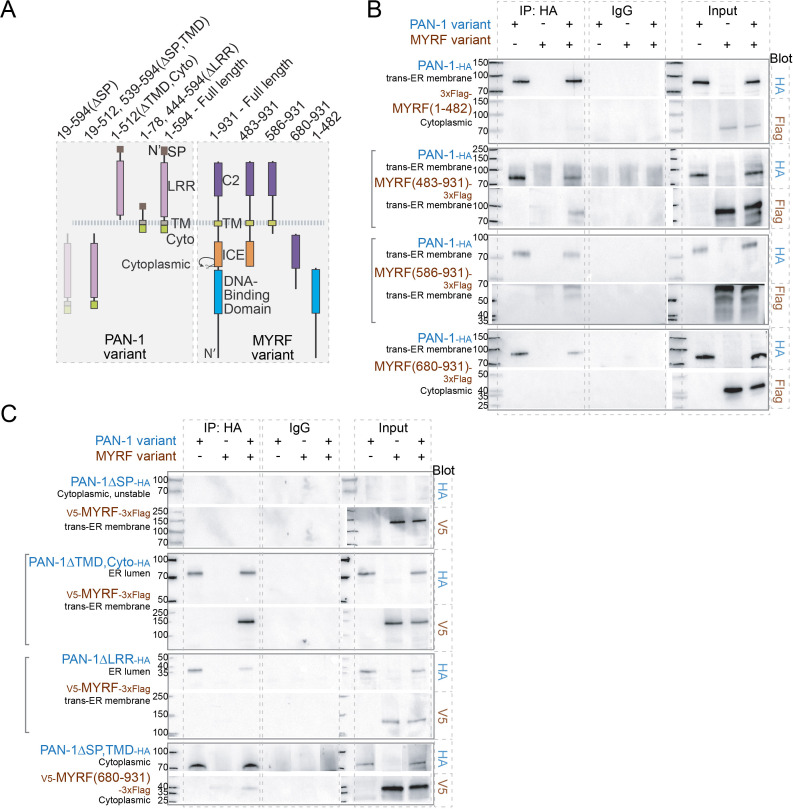
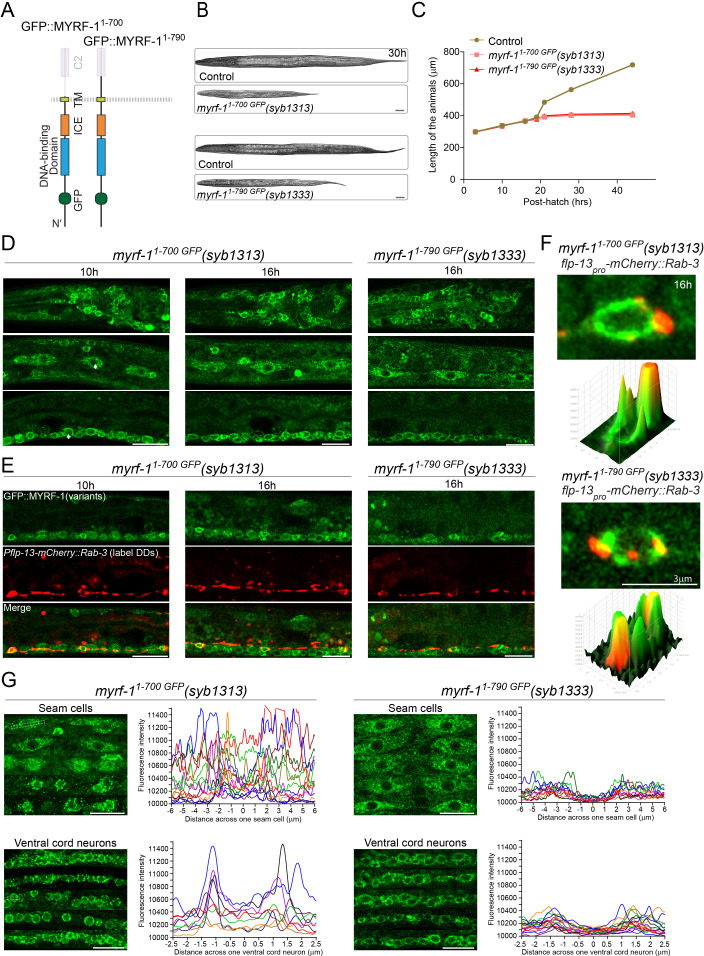
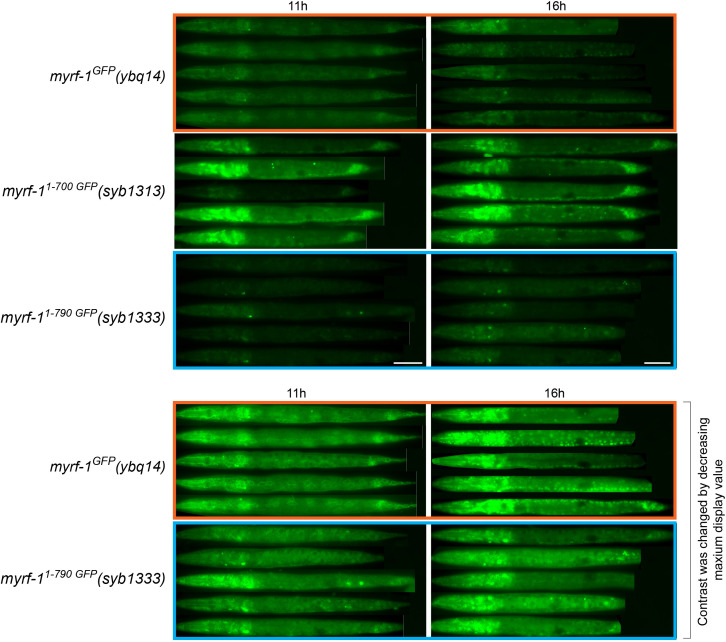
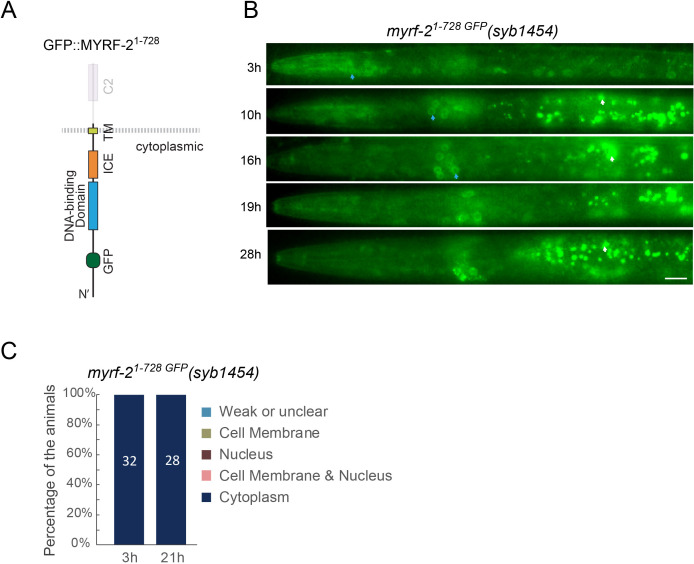
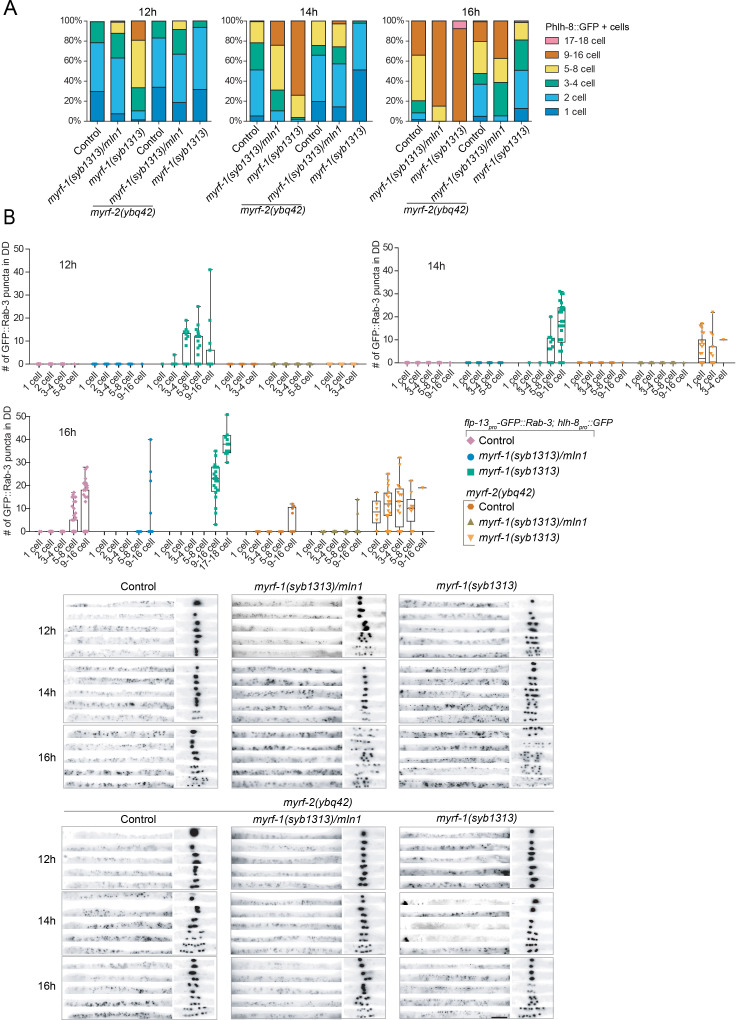
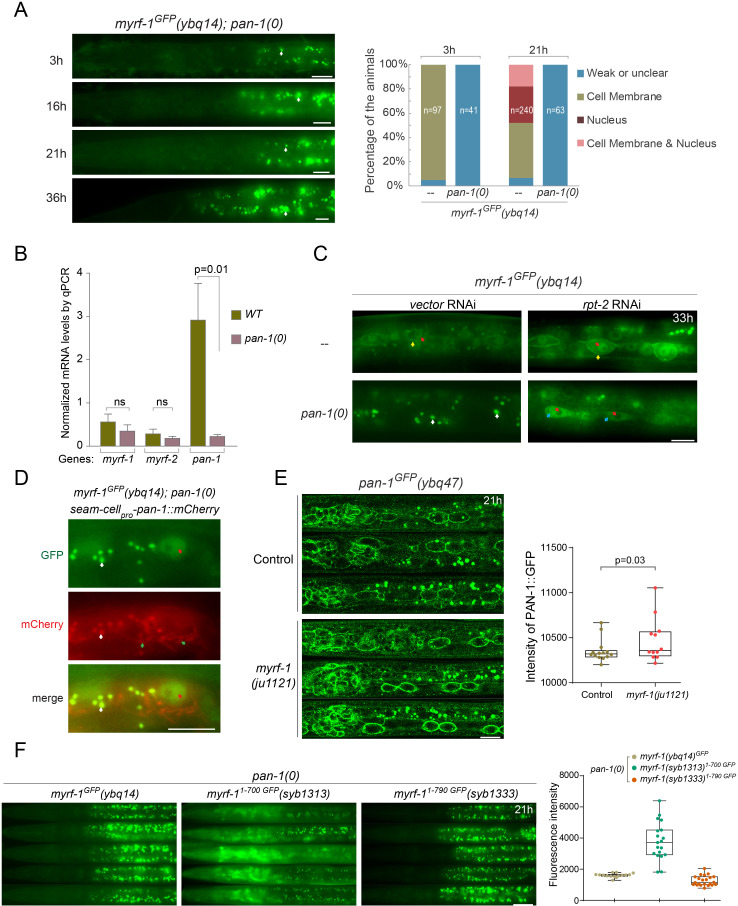
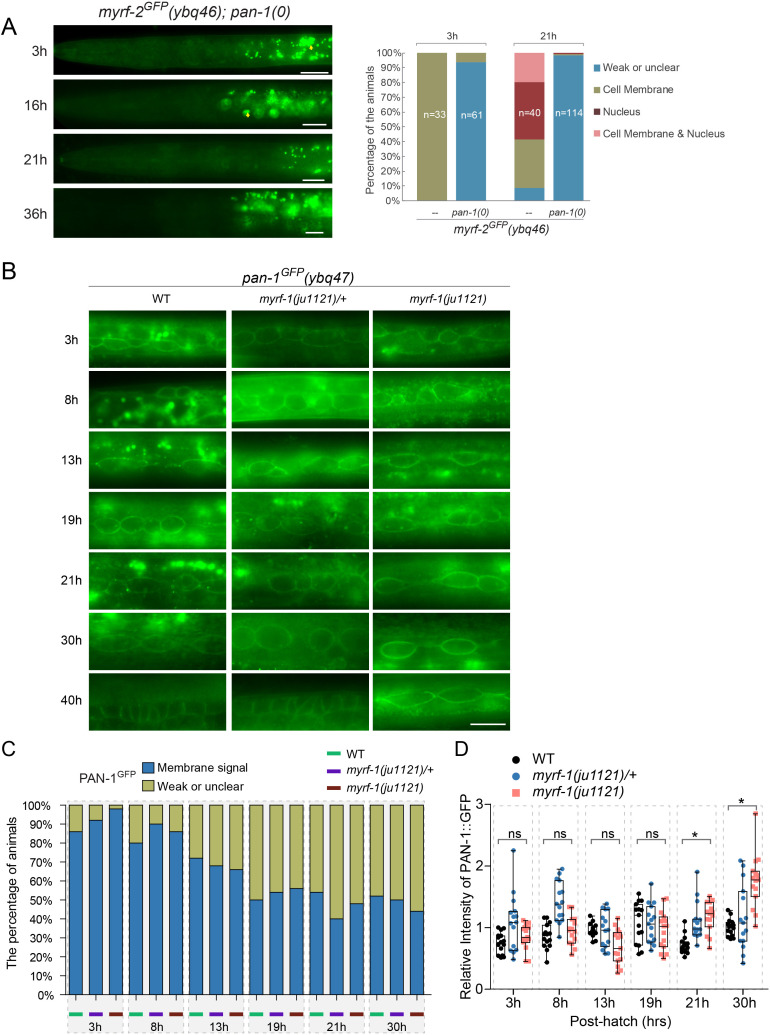
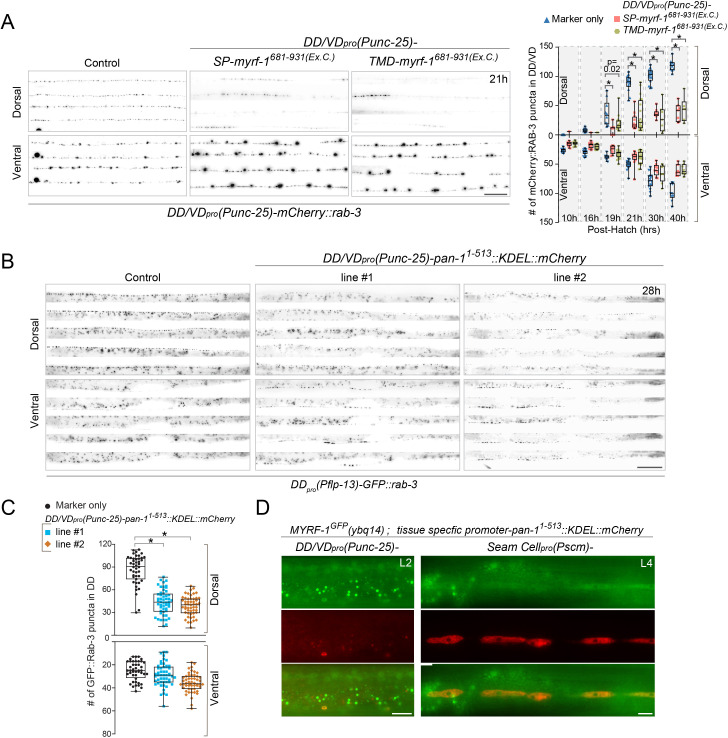
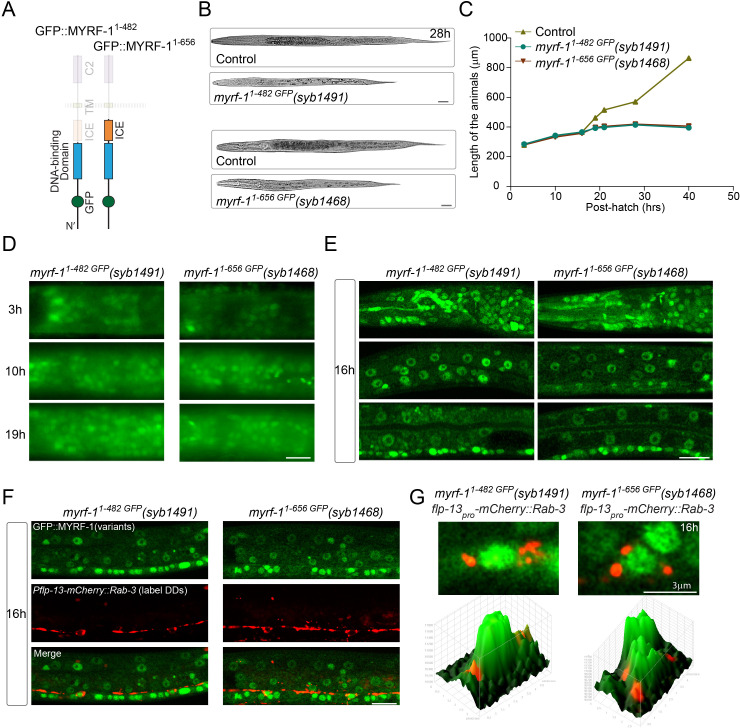
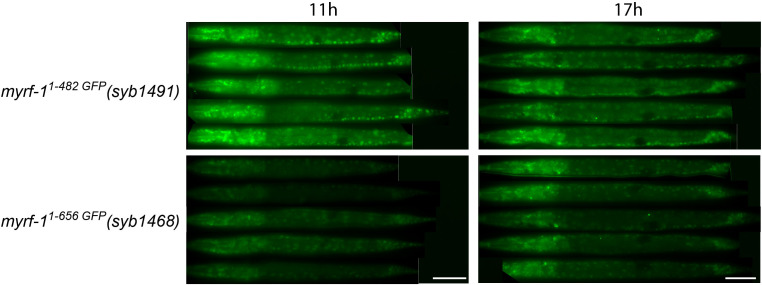
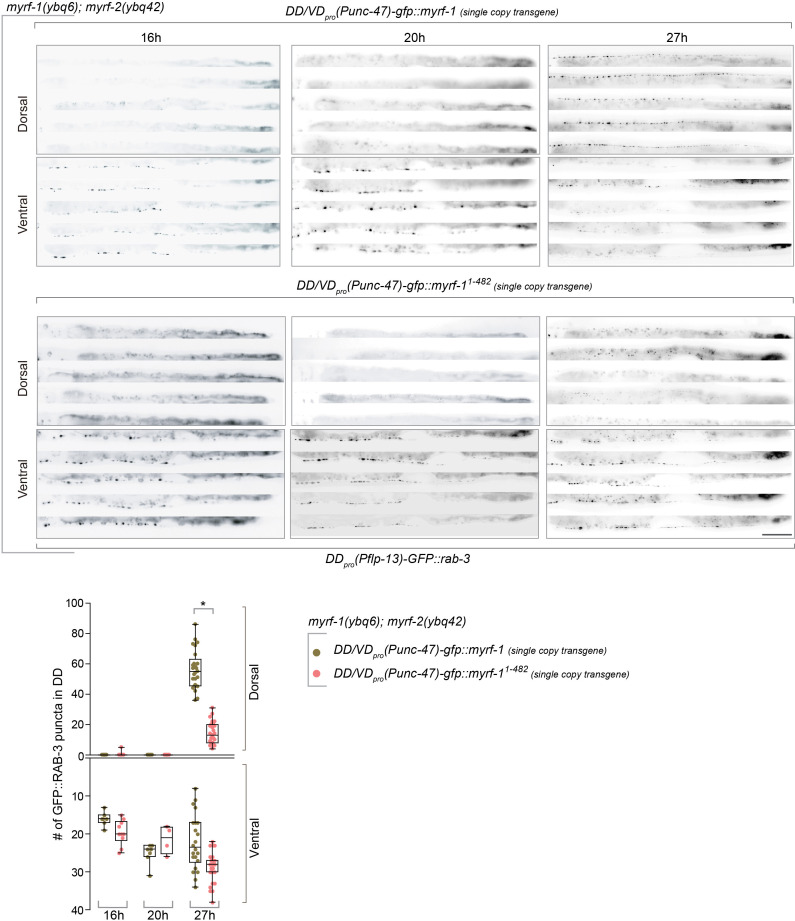
Neural circuits develop through a plastic phase orchestrated by genetic programs and environmental signals. We have identified a leucine-rich-repeat domain transmembrane protein PAN-1 as a factor required for synaptic rewiring in . PAN-1 localizes on cell membrane and binds with MYRF, a membrane-bound transcription factor indispensable for promoting synaptic rewiring. Full-length MYRF was known to undergo self-cleavage on ER membrane and release its transcriptional N-terminal fragment in cultured cells. We surprisingly find that MYRF trafficking to cell membrane before cleavage is pivotal for development and the timing of N-MYRF release coincides with the onset of synaptic rewiring. On cell membrane PAN-1 and MYRF interact with each other via their extracellular regions. Loss of PAN-1 abolishes MYRF cell membrane localization, consequently blocking -dependent neuronal rewiring process. Thus, through interactions with a cooperating factor on the cell membrane, MYRF may link cell surface activities to transcriptional cascades required for development.
神经回路通过遗传程序和环境信号协调的可塑性阶段来发育。我们已经确定了富含亮氨酸的重复域跨膜蛋白 PAN-1 作为突触重连所必需的因素。PAN-1 定位于细胞膜上,并与 MYRF 结合,MYRF 是一种膜结合转录因子,对于促进突触重连是必不可少的。全长 MYRF 已知在 ER 膜上发生自我切割,并在培养细胞中释放其转录的 N 端片段。我们惊讶地发现,MYRF 在切割前向细胞膜的运输对于发育至关重要,并且 N-MYRF 的释放时间与突触重连的开始时间一致。在细胞膜上,PAN-1 和 MYRF 通过其细胞外区域相互作用。PAN-1 的缺失会破坏 MYRF 的细胞膜定位,从而阻止 -dependent 神经元重连过程。因此,通过与细胞膜上的合作因子相互作用,MYRF 可能将细胞表面活性与发育所需的转录级联联系起来。