Weill Institute for Cell and Molecular Biology, Cornell University, Ithaca, United States.
Department of Molecular Biology and Genetics, Cornell University, Ithaca, United States.
Elife. 2021 May 24;10:e67709. doi: 10.7554/eLife.67709.
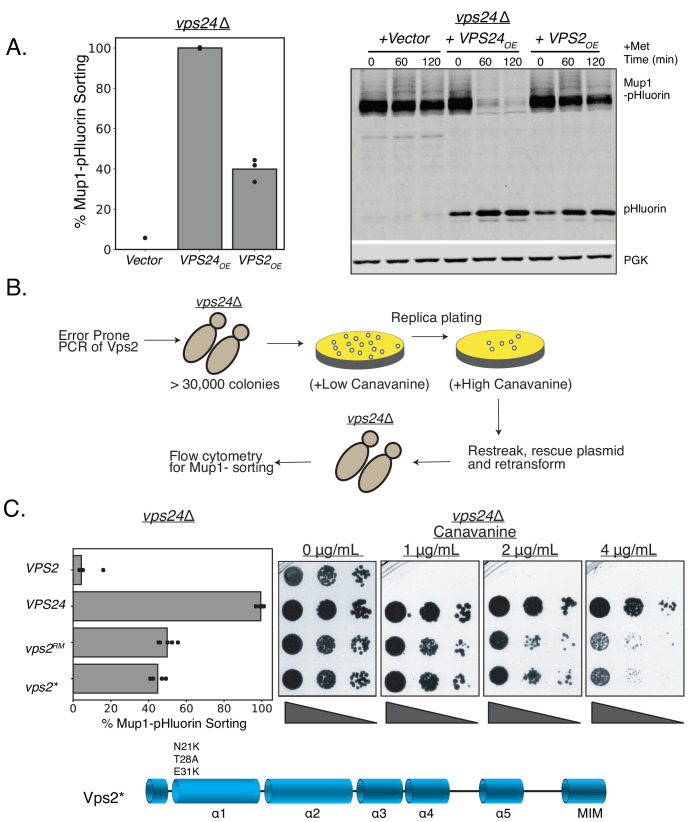
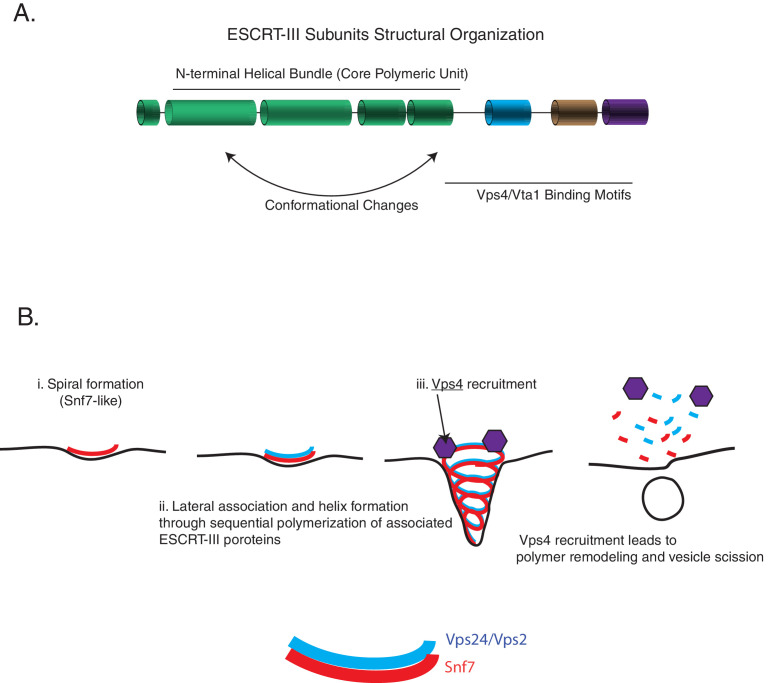
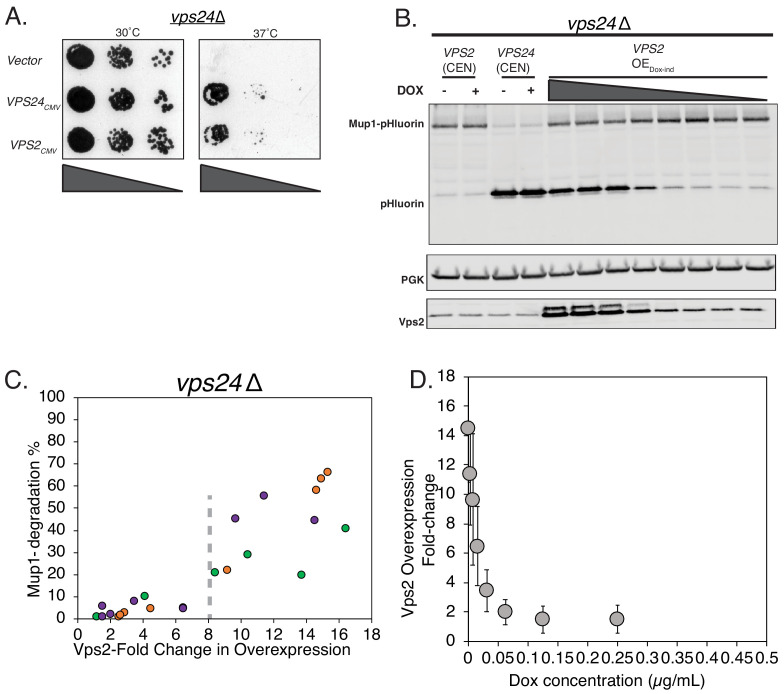
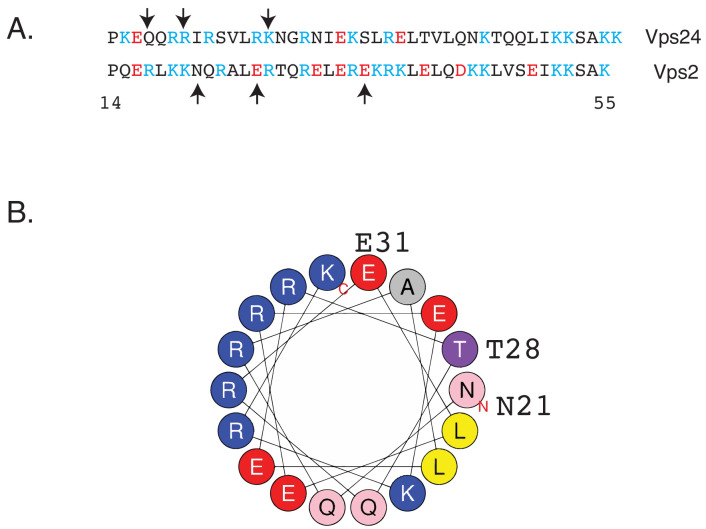
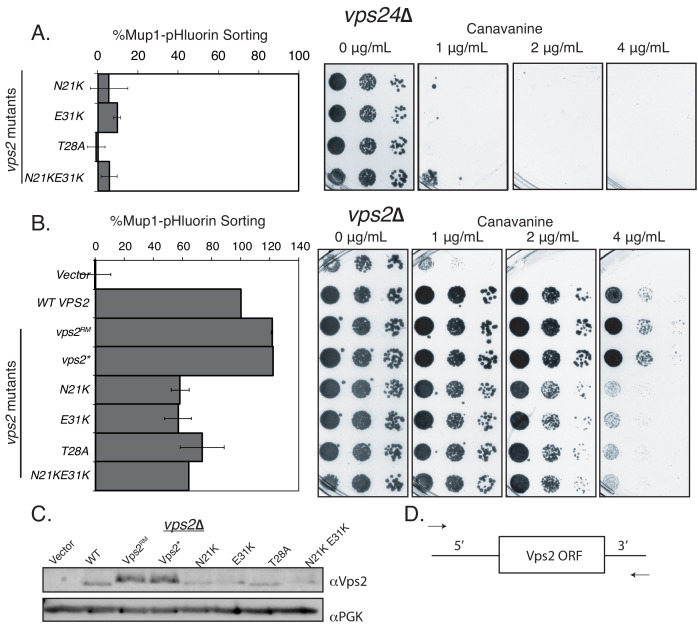
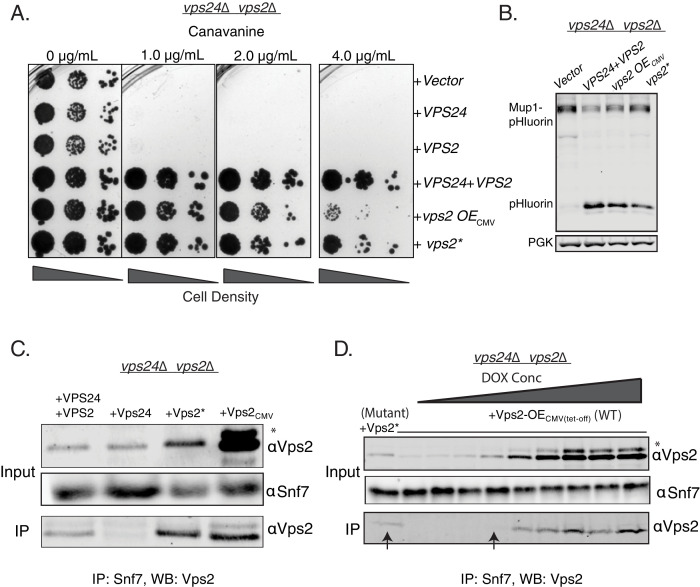
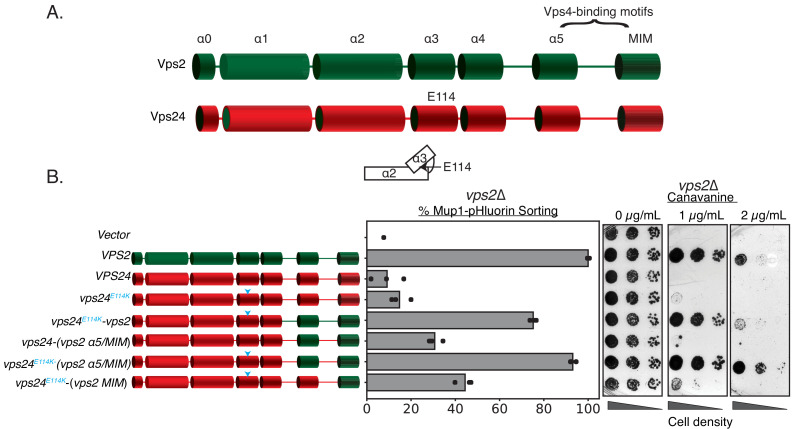
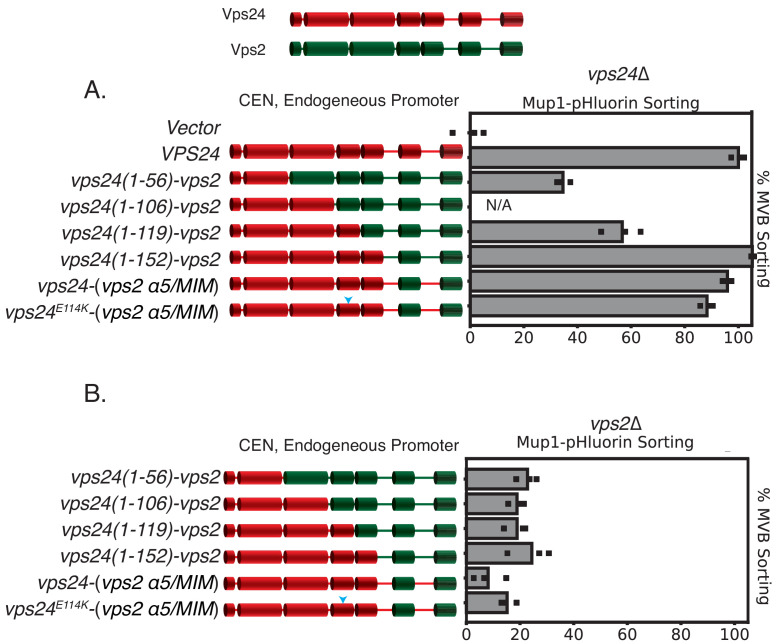
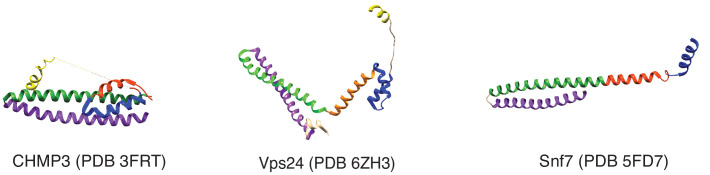
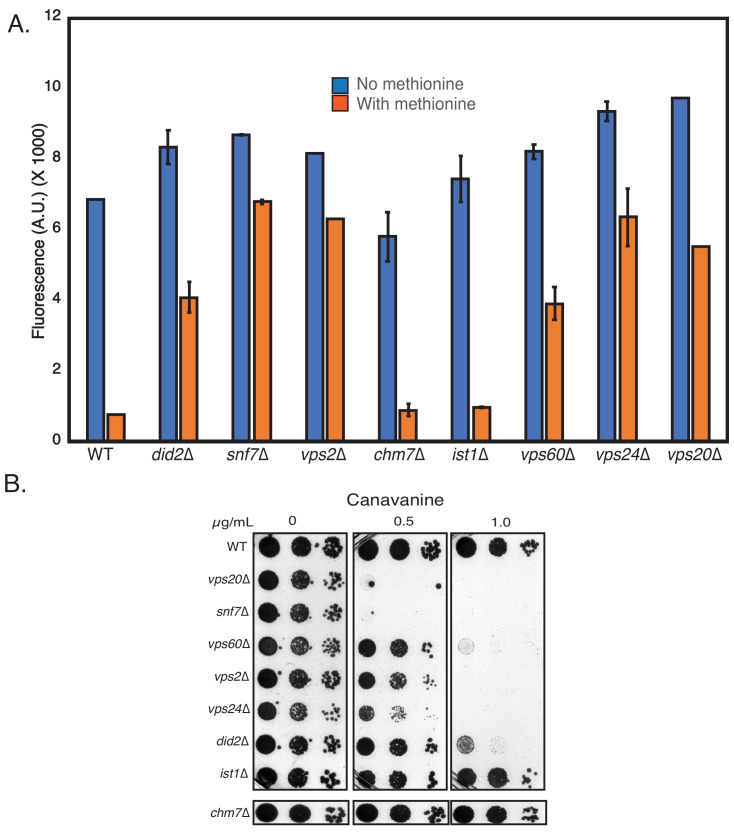
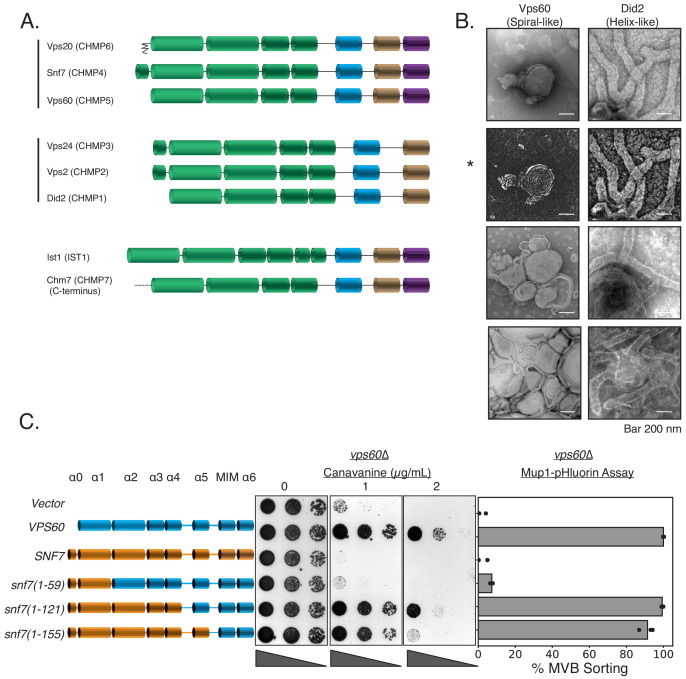
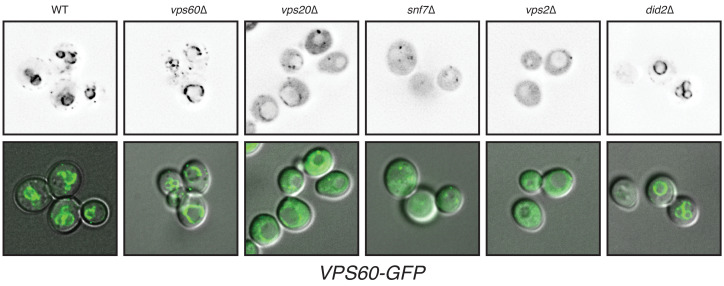
ESCRT-III polymerization is required for all endosomal sorting complex required for transport (ESCRT)-dependent events in the cell. However, the relative contributions of the eight ESCRT-III subunits differ between each process. The minimal features of ESCRT-III proteins necessary for function and the role for the multiple ESCRT-III subunits remain unclear. To identify essential features of ESCRT-III subunits, we previously studied the polymerization mechanisms of two ESCRT-III subunits Snf7 and Vps24, identifying the association of the helix-4 region of Snf7 with the helix-1 region of Vps24 (Banjade et al., 2019a). Here, we find that mutations in the helix-1 region of another ESCRT-III subunit Vps2 can functionally replace Vps24 in . Engineering and genetic selections revealed the required features of both subunits. Our data allow us to propose three minimal features required for ESCRT-III function - spiral formation, lateral association of the spirals through heteropolymerization, and binding to the AAA + ATPase Vps4 for dynamic remodeling.
ESCRT-III 聚合对于细胞内所有依赖于内体分选复合物(ESCRT)的运输事件都是必需的。然而,每个过程中八个 ESCRT-III 亚基的相对贡献不同。ESCRT-III 蛋白发挥功能的最小特征以及多个 ESCRT-III 亚基的作用仍不清楚。为了确定 ESCRT-III 亚基的必需特征,我们之前研究了两个 ESCRT-III 亚基 Snf7 和 Vps24 的聚合机制,确定了 Snf7 的螺旋 4 区与 Vps24 的螺旋 1 区的结合(Banjade 等人,2019a)。在这里,我们发现另一个 ESCRT-III 亚基 Vps2 的螺旋 1 区的突变可以在功能上取代 Vps24。工程和遗传选择揭示了两个亚基所需的特征。我们的数据使我们能够提出 ESCRT-III 功能所需的三个最小特征-螺旋形成、通过异聚化横向连接螺旋、以及与 AAA+ATPase Vps4 结合进行动态重塑。