Latvian Biomedicine Research and Study Centre, LV-1067 Riga, Latvia.
R.E. Kavetsky Institute of Experimental Pathology, Oncology and Radiobiology, National Academy of Sciences of Ukraine, 03022 Kyiv, Ukraine.
Cells. 2021 Jun 23;10(7):1582. doi: 10.3390/cells10071582.
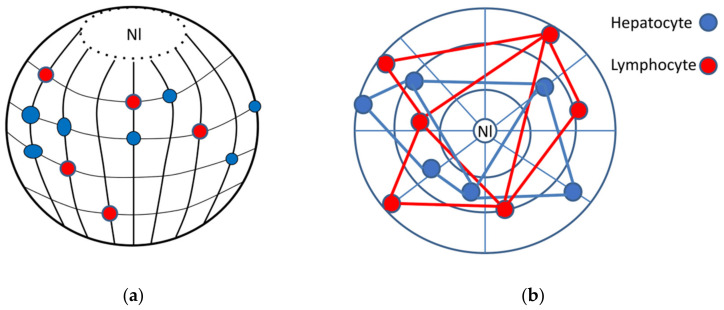
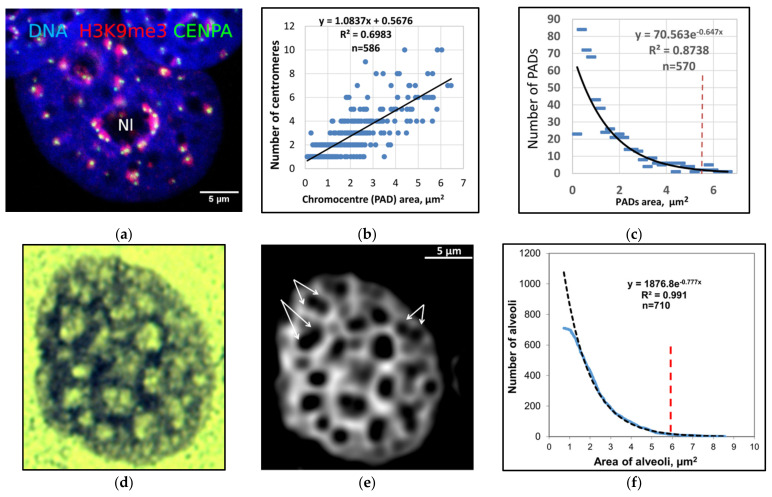
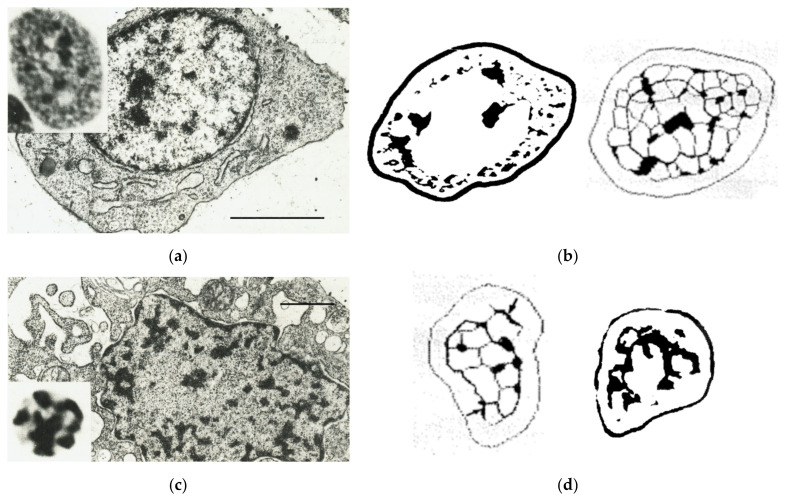
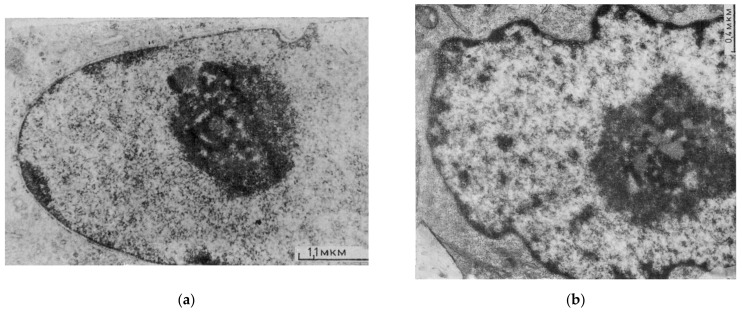
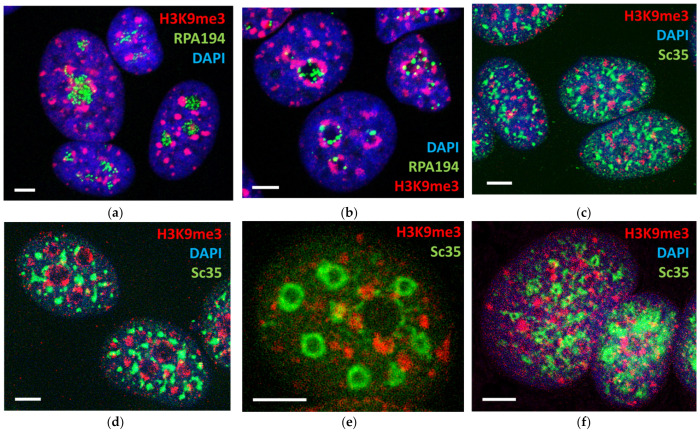
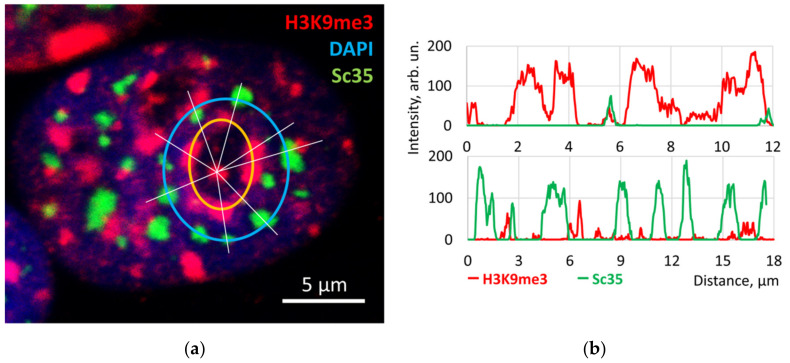
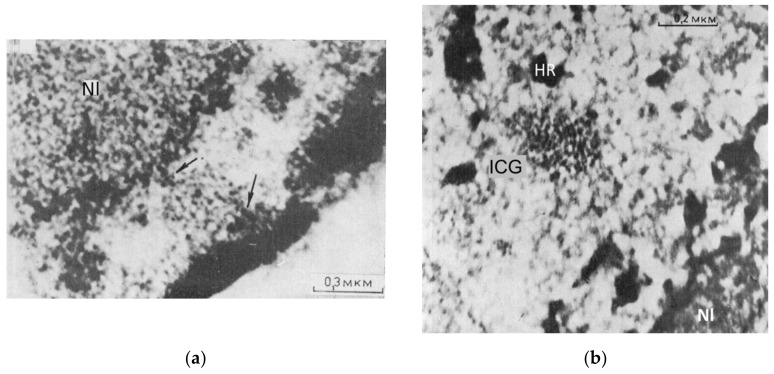
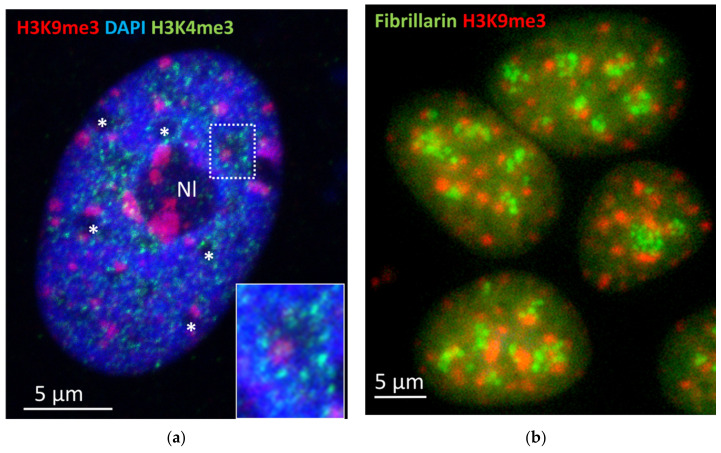
Open systems can only exist by self-organization as pulsing structures exchanging matter and energy with the outer world. This review is an attempt to reveal the organizational principles of the heterochromatin supra-intra-chromosomal network in terms of nonlinear thermodynamics. The accessibility of the linear information of the genetic code is regulated by constitutive heterochromatin (CHR) creating the positional information in a system of coordinates. These features include scale-free splitting-fusing of CHR with the boundary constraints of the nucleolus and nuclear envelope. The analysis of both the literature and our own data suggests a radial-concentric network as the main structural organization principle of CHR regulating transcriptional pulsing. The dynamic CHR network is likely created together with nucleolus-associated chromatin domains, while the alveoli of this network, including springy splicing speckles, are the pulsing transcription hubs. CHR contributes to this regulation due to the silencing position variegation effect, stickiness, and flexible rigidity determined by the positioning of nucleosomes. The whole system acts in concert with the elastic nuclear actomyosin network which also emerges by self-organization during the transcriptional pulsing process. We hypothesize that the the transcriptional pulsing, in turn, adjusts its frequency/amplitudes specified by topologically associating domains to the replication timing code that determines epigenetic differentiation memory.
开放系统只能通过自组织作为脉冲结构与外部世界交换物质和能量而存在。本综述试图从非线性热力学的角度揭示异染色质超染色体网络的组织原则。线性遗传密码信息的可及性受组成型异染色质(CHR)调节,在坐标系中创建位置信息。这些特征包括无标度的 CHR 分裂-融合,边界约束核仁核膜。对文献和我们自己数据的分析表明,径向同心网络是调节转录脉冲的 CHR 的主要结构组织原则。动态 CHR 网络可能与核仁相关染色质域一起创建,而该网络的肺泡,包括有弹性的剪接斑点,是脉冲转录枢纽。CHR 通过由核小体定位决定的沉默位置变异效应、粘性和弹性刚性对这种调节做出贡献。整个系统与弹性核肌动球蛋白网络协同作用,弹性核肌动球蛋白网络也在转录脉冲过程中通过自组织产生。我们假设,转录脉冲反过来又会根据拓扑关联域调整其频率/幅度指定的复制时间代码,以确定决定表观遗传分化记忆的复制时间代码。