Liu Li, Schubert Daniel M, Könneke Martin, Berg Ivan A
Institute for Molecular Microbiology and Biotechnology, University of Münster, Münster, Germany.
Department of Microbiology, Faculty of Biology, University of Freiburg, Freiburg, Germany.
Front Microbiol. 2021 Jul 5;12:712030. doi: 10.3389/fmicb.2021.712030. eCollection 2021.
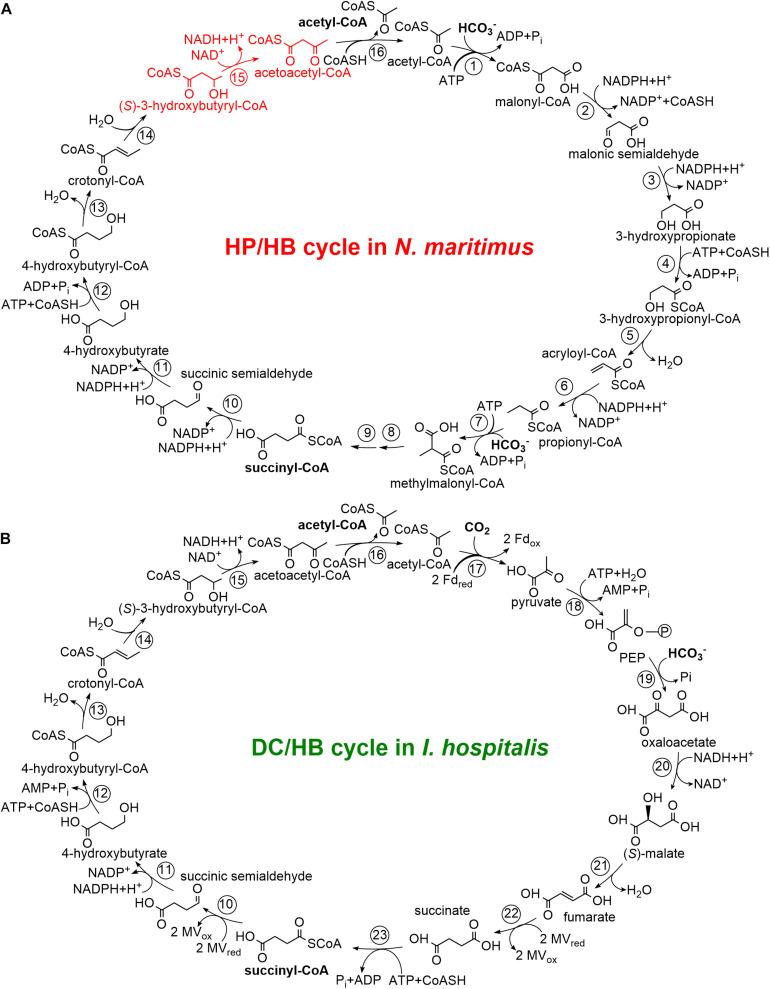
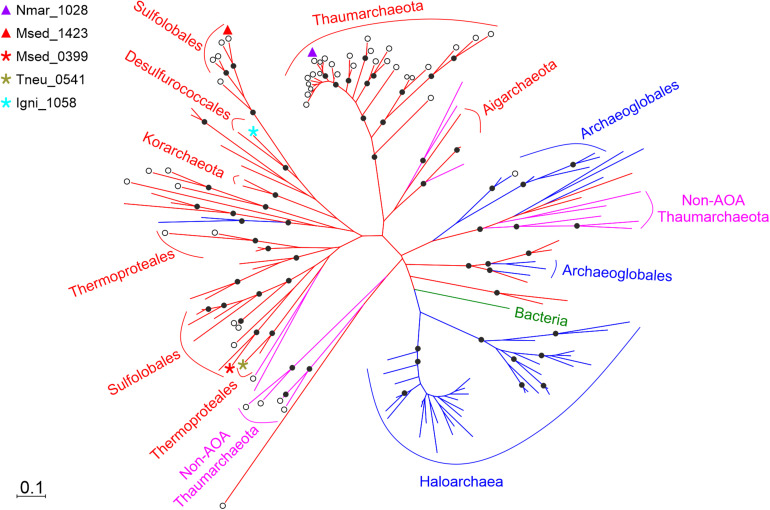
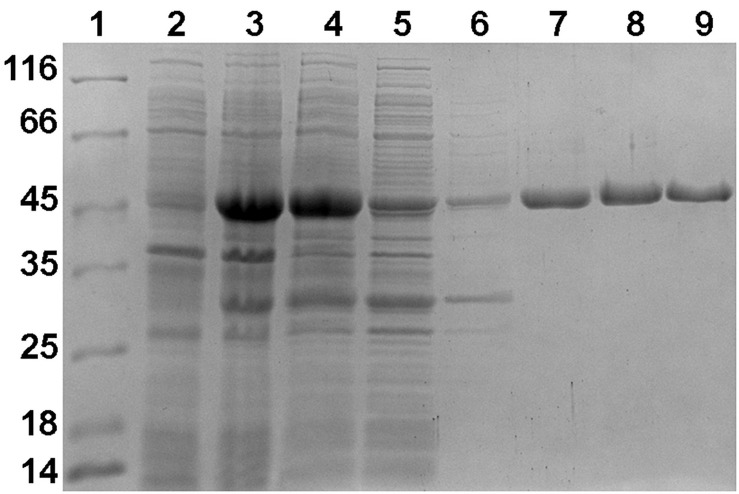
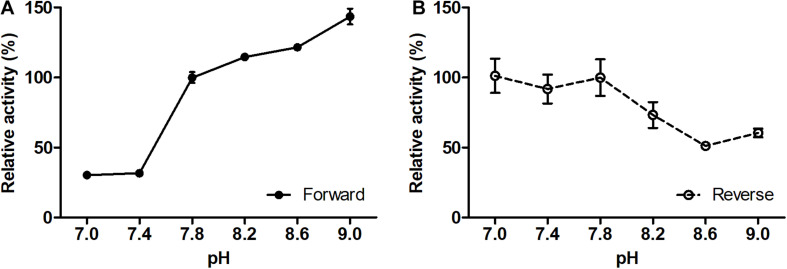
Ammonia-oxidizing archaea of the phylum Thaumarchaeota are among the most abundant organisms that exert primary control of oceanic and soil nitrification and are responsible for a large part of dark ocean primary production. They assimilate inorganic carbon via an energetically efficient version of the 3-hydroxypropionate/4-hydroxybutyrate cycle. In this cycle, acetyl-CoA is carboxylated to succinyl-CoA, which is then converted to two acetyl-CoA molecules with 4-hydroxybutyrate as the key intermediate. This conversion includes the ()-3-hydroxybutyryl-CoA dehydrogenase reaction. Here, we heterologously produced the protein Nmar_1028 catalyzing this reaction in thaumarchaeon , characterized it biochemically and performed its phylogenetic analysis. This NAD-dependent dehydrogenase is highly active with its substrate, ()-3-hydroxybutyryl-CoA, and its low value suggests that the protein is adapted to the functioning in the 3-hydroxypropionate/4-hydroxybutyrate cycle. Nmar_1028 is homologous to the dehydrogenase domain of crotonyl-CoA hydratase/()-3-hydroxybutyryl-CoA dehydrogenase that is present in many Archaea. Apparently, the loss of the dehydratase domain of the fusion protein in the course of evolution was accompanied by lateral gene transfer of 3-hydroxypropionyl-CoA dehydratase/crotonyl-CoA hydratase from Bacteria. Although ()-3-hydroxybutyryl-CoA dehydrogenase studied here is neither unique nor characteristic for the HP/HB cycle, Nmar_1028 appears to be the only ()-3-hydroxybutyryl-CoA dehydrogenase in and is thus essential for the functioning of the 3-hydroxypropionate/4-hydroxybutyrate cycle and for the biology of this important marine archaeon.
泉古菌门的氨氧化古菌是对海洋和土壤硝化作用施加主要控制的最丰富生物之一,并且在很大程度上负责海洋暗区初级生产。它们通过3-羟基丙酸酯/4-羟基丁酸酯循环的高效能量版本同化无机碳。在这个循环中,乙酰辅酶A羧化生成琥珀酰辅酶A,然后以4-羟基丁酸作为关键中间体将其转化为两个乙酰辅酶A分子。这种转化包括()-3-羟基丁酰辅酶A脱氢酶反应。在这里,我们在泉古菌中异源表达催化该反应的蛋白质Nmar_1028,对其进行生化表征并进行系统发育分析。这种依赖NAD的脱氢酶对其底物()-3-羟基丁酰辅酶A具有高活性,其低 值表明该蛋白质适合在3-羟基丙酸酯/4-羟基丁酸酯循环中发挥作用。Nmar_1028与许多古菌中存在的巴豆酰辅酶A水合酶/()-3-羟基丁酰辅酶A脱氢酶的脱氢酶结构域同源。显然,在进化过程中融合蛋白脱水酶结构域的丧失伴随着来自细菌的3-羟基丙酰辅酶A脱水酶/巴豆酰辅酶A水合酶的横向基因转移。尽管这里研究的()-3-羟基丁酰辅酶A脱氢酶对于HP/HB循环既不是独特的也不是其特征性的,但Nmar_1028似乎是泉古菌中唯一的()-3-羟基丁酰辅酶A脱氢酶,因此对于3-羟基丙酸酯/4-羟基丁酸酯循环的功能以及这种重要海洋古菌的生物学特性至关重要。