Biomedical Center (BMC), Divison of Molecular Biology, Faculty of Medicine, LMU Munich, 82152 Munich, Germany.
Int J Mol Sci. 2021 Jul 30;22(15):8233. doi: 10.3390/ijms22158233.
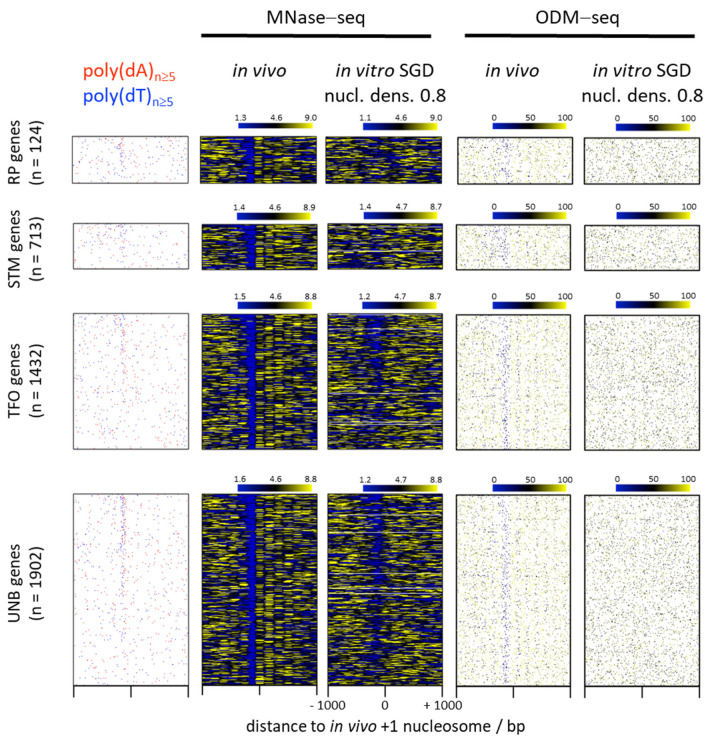
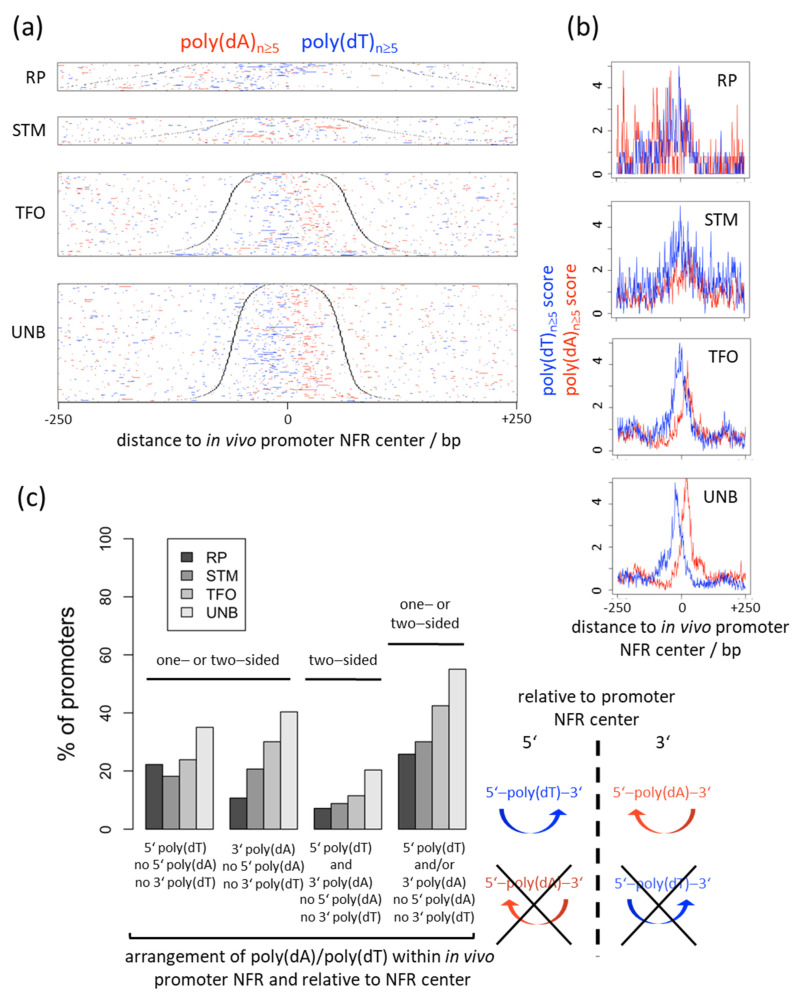
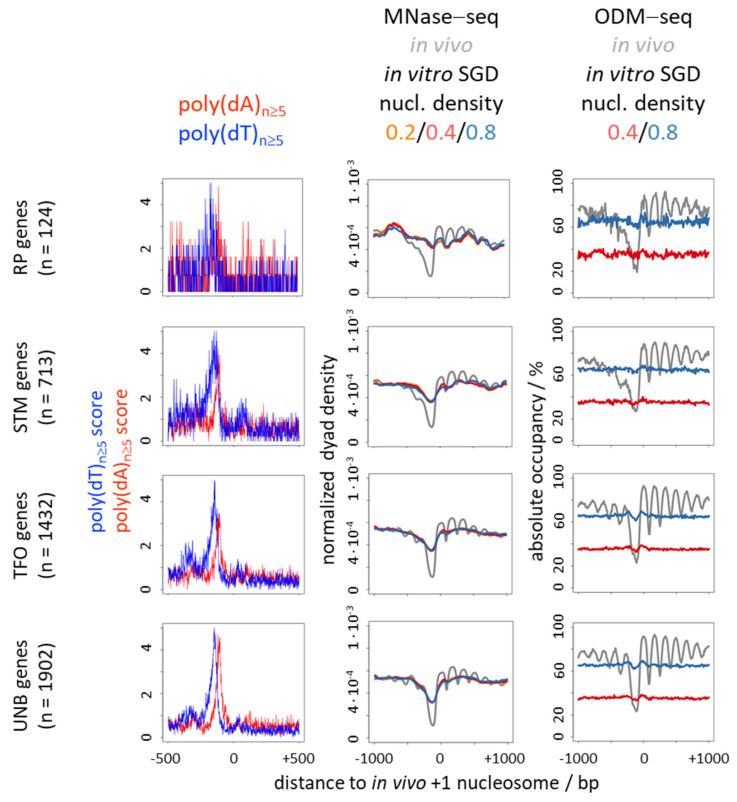
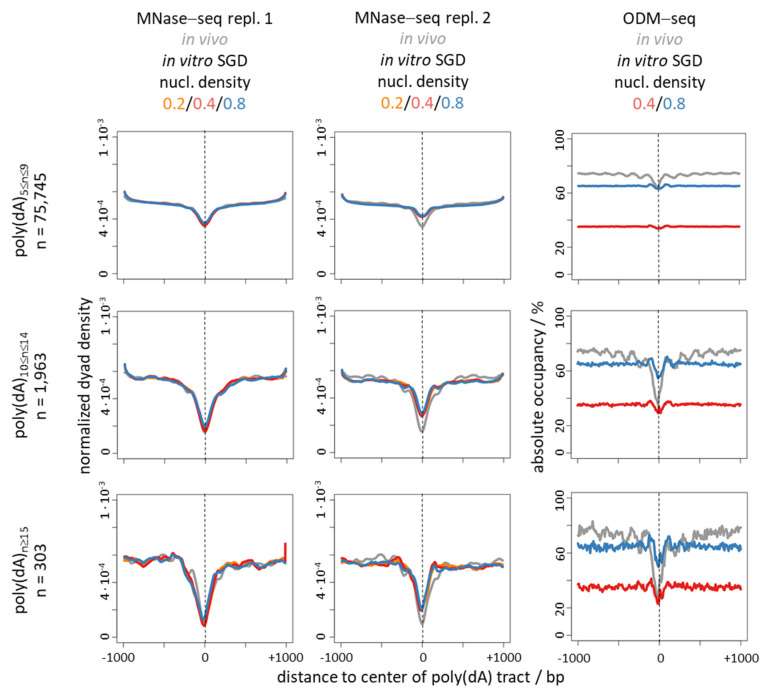
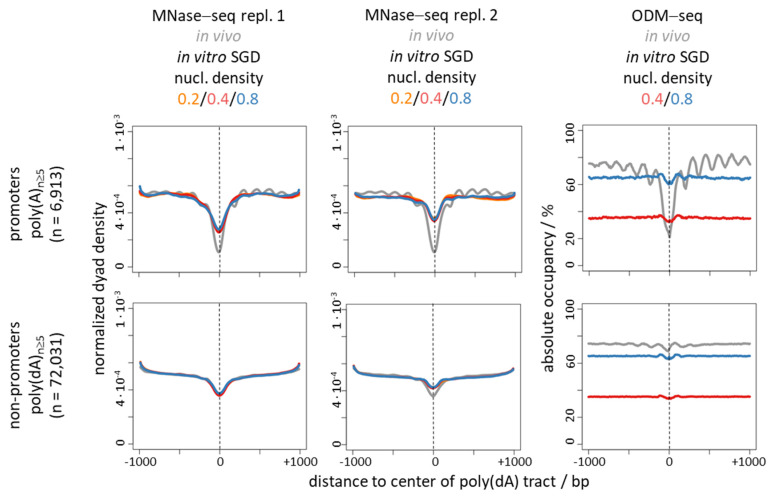
Poly(dA:dT) tracts cause nucleosome depletion in many species, e.g., at promoters and replication origins. Their intrinsic biophysical sequence properties make them stiff and unfavorable for nucleosome assembly, as probed by in vitro nucleosome reconstitution. The mere correlation between nucleosome depletion over poly(dA:dT) tracts in in vitro reconstituted and in in vivo chromatin inspired an intrinsic nucleosome exclusion mechanism in vivo that is based only on DNA and histone properties. However, we compile here published and new evidence that this correlation does not reflect mechanistic causation. (1) Nucleosome depletion over poly(dA:dT) in vivo is not universal, e.g., very weak in . (2) The energy penalty for incorporating poly(dA:dT) tracts into nucleosomes is modest (<10%) relative to ATP hydrolysis energy abundantly invested by chromatin remodelers. (3) Nucleosome depletion over poly(dA:dT) is much stronger in vivo than in vitro if monitored without MNase and (4) actively maintained in vivo. (5) promoters evolved a strand-biased poly(dA) versus poly(dT) distribution. (6) Nucleosome depletion over poly(dA) is directional in vivo. (7) The ATP dependent chromatin remodeler RSC preferentially and directionally displaces nucleosomes towards 5' of poly(dA). Especially distribution strand bias and displacement directionality would not be expected for an intrinsic mechanism. Together, this argues for an in vivo mechanism where active and species-specific read out of intrinsic sequence properties, e.g., by remodelers, shapes nucleosome organization.
多聚(dA:dT)序列在许多物种中导致核小体耗竭,例如在启动子和复制起点处。它们的固有生物物理序列特性使它们变得僵硬,不利于核小体组装,这可以通过体外核小体重建来探测。仅仅基于体外重建和体内染色质中多聚(dA:dT)序列上核小体耗竭的相关性,就启发了一种仅基于 DNA 和组蛋白特性的体内固有核小体排除机制。然而,我们在这里汇编了已发表和新的证据,表明这种相关性并不反映机制因果关系。(1)体内多聚(dA:dT)上的核小体耗竭并非普遍存在,例如在. 中非常微弱。(2)将多聚(dA:dT)序列纳入核小体的能量代价相对染色质重塑剂大量投入的 ATP 水解能量来说是适度的(<10%)。(3)如果不使用 MNase 监测,多聚(dA:dT)上的核小体耗竭在体内比在体外要强得多,并且(4)在体内被积极维持。(5)启动子进化出了链偏倚的多聚(dA)与多聚(dT)分布。(6)体内核小体耗竭具有方向性。(7)ATP 依赖的染色质重塑器 RSC 优先且有方向性地将核小体向多聚(dA)的 5'方向位移。特别是分布链偏倚和位移方向性对于内在机制是不可预期的。总之,这表明存在一种体内机制,其中活跃且具有物种特异性的内在序列特性的读取,例如通过重塑器,塑造核小体组织。