Department of Molecular and Cellular Biology, Harvard University, Cambridge, United States.
Department of Organismic and Evolutionary Biology, Harvard University, Cambridge, United States.
Elife. 2021 Oct 1;10:e70918. doi: 10.7554/eLife.70918.
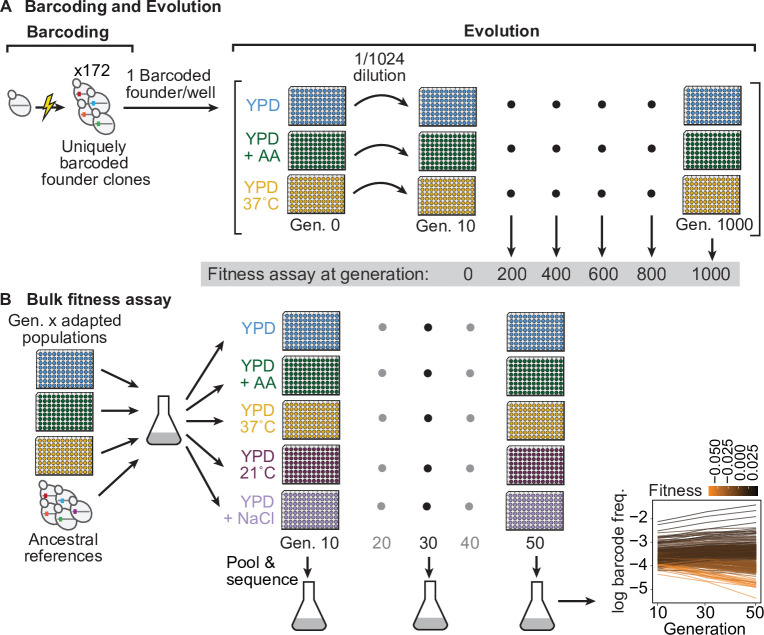
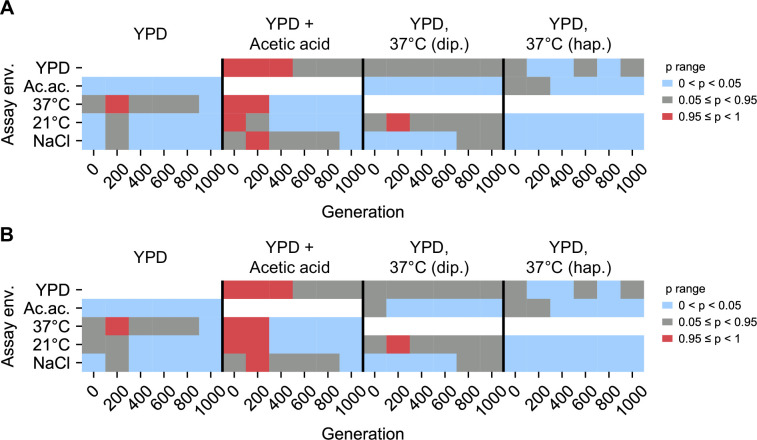
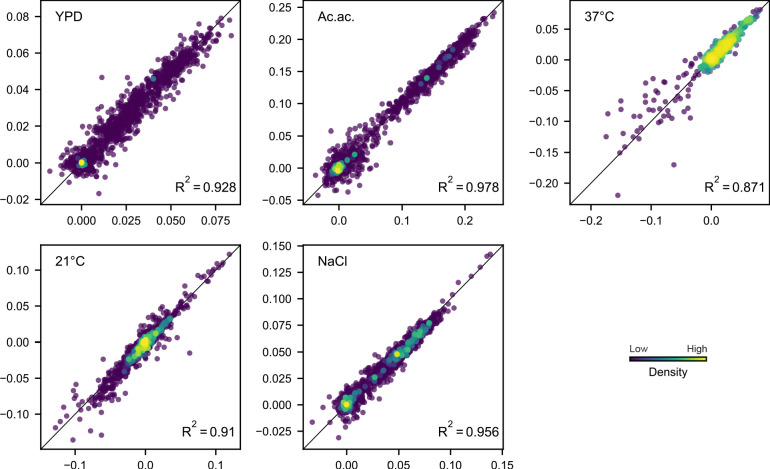
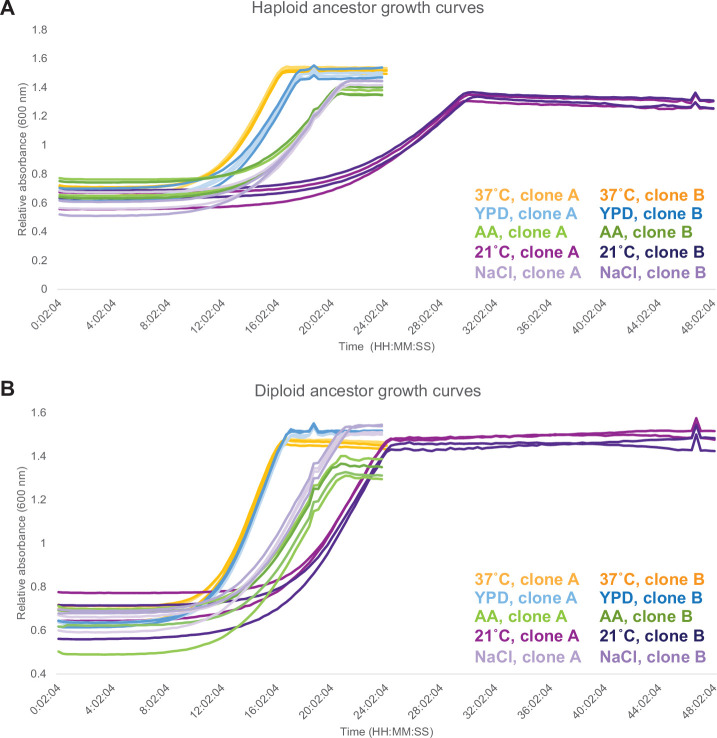
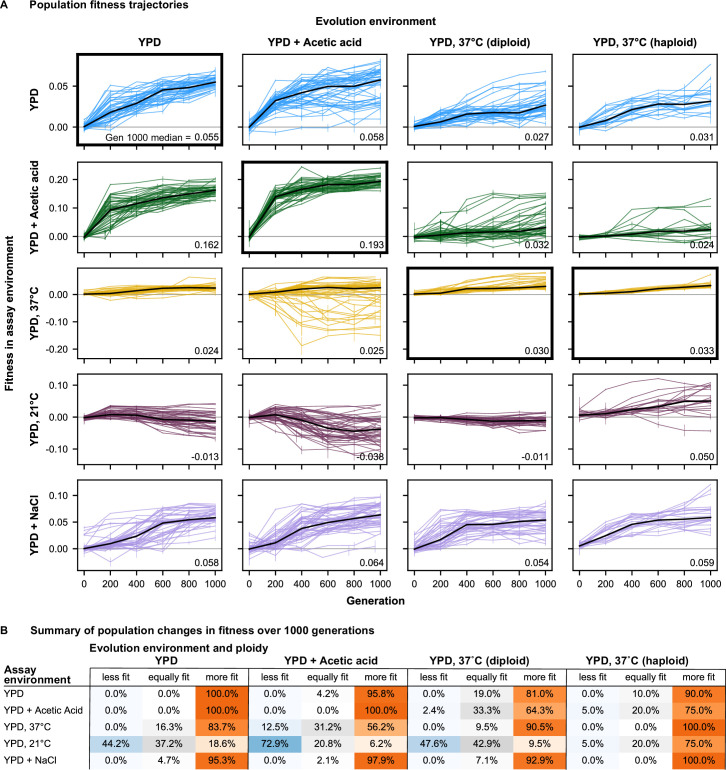
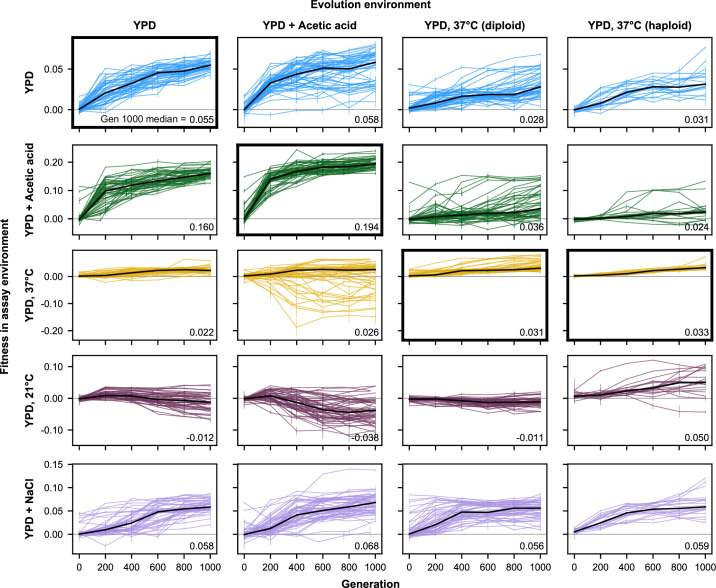
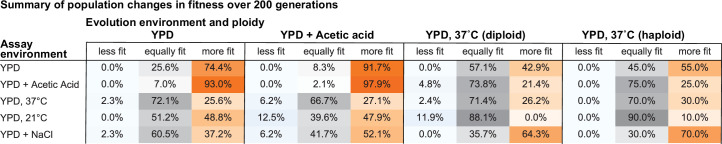
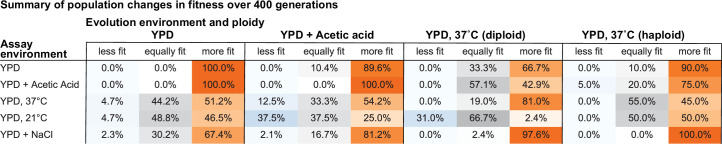
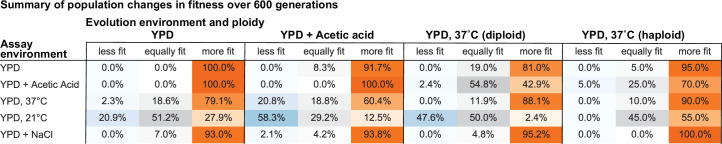
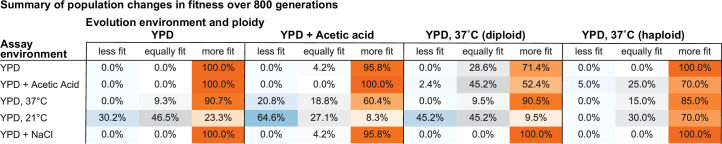
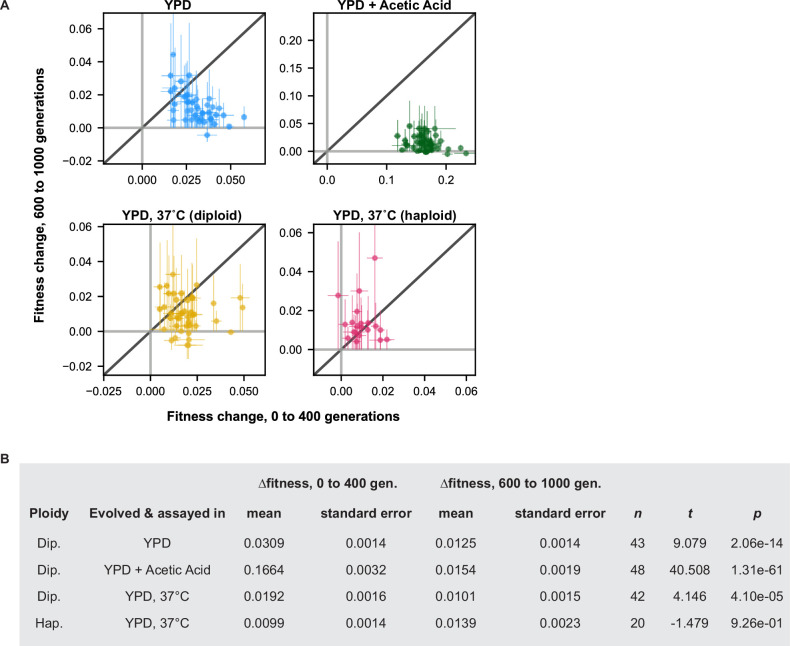
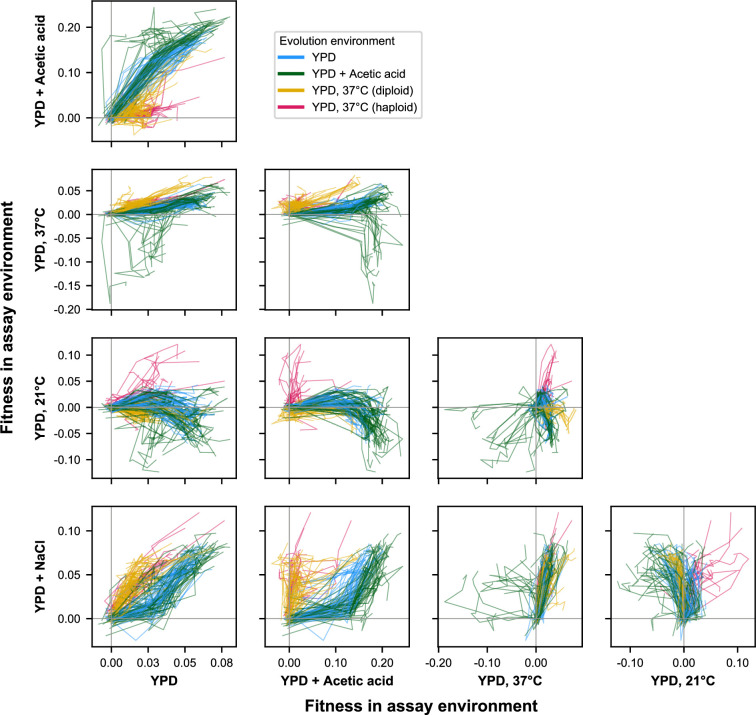
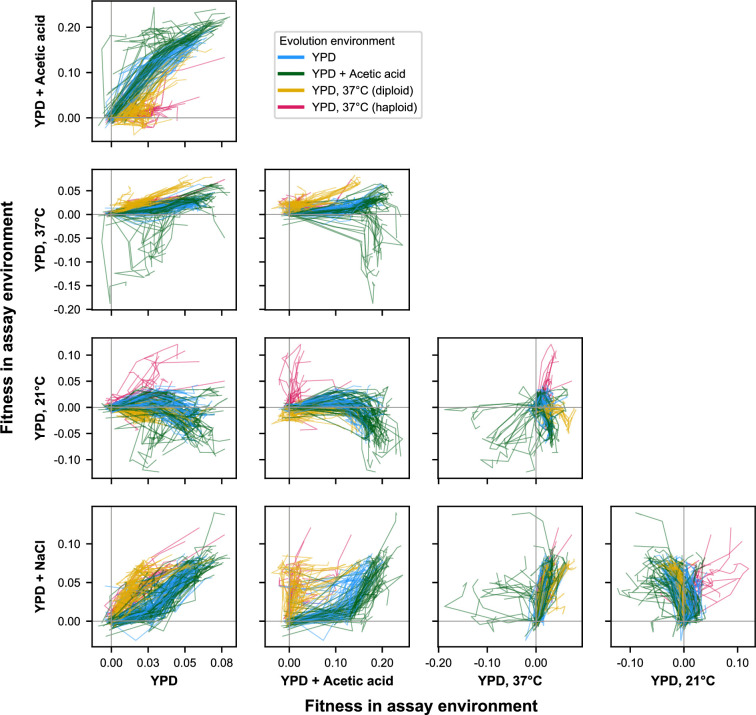
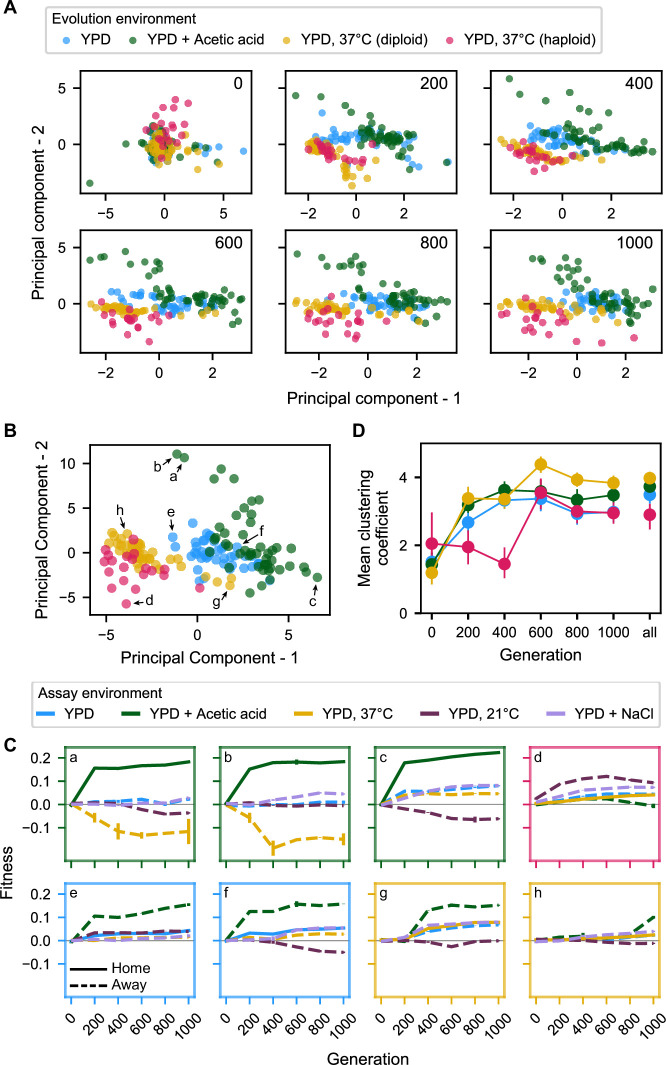
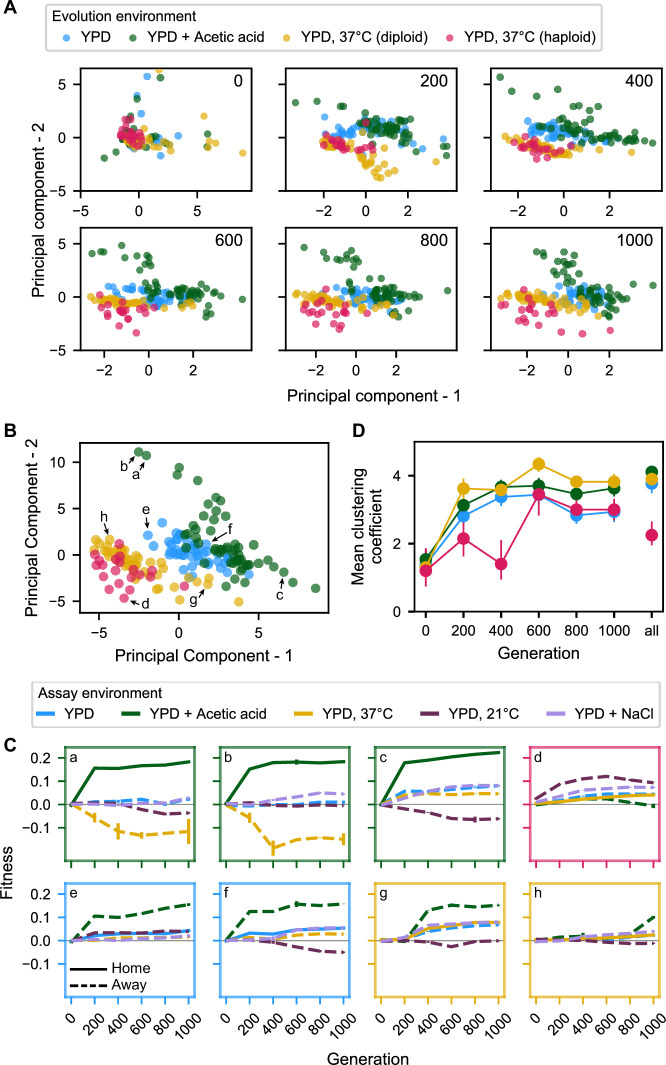
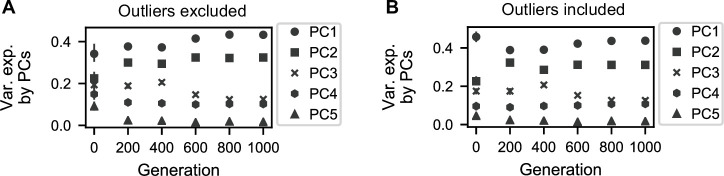
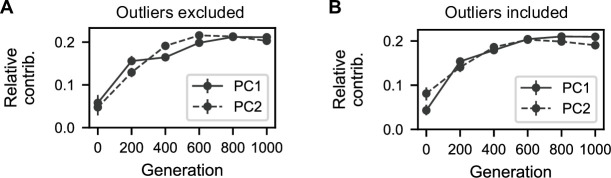
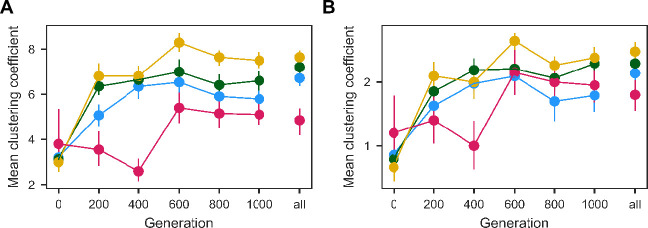
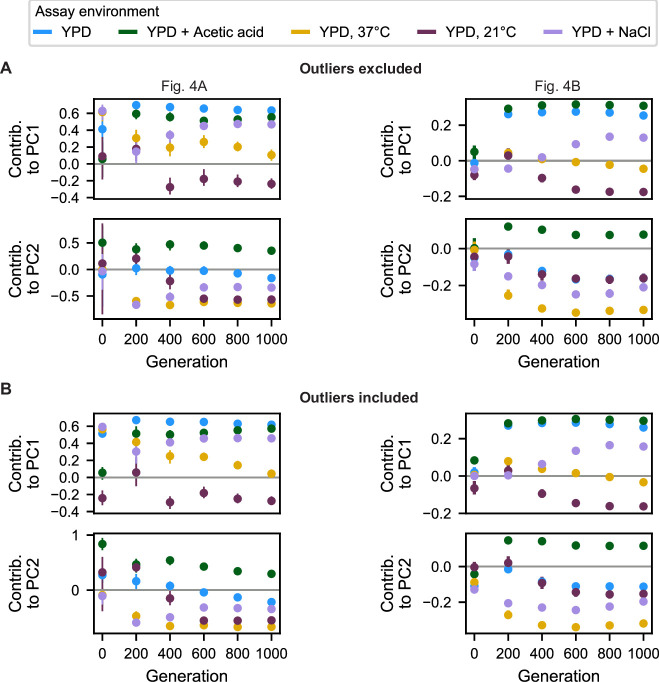
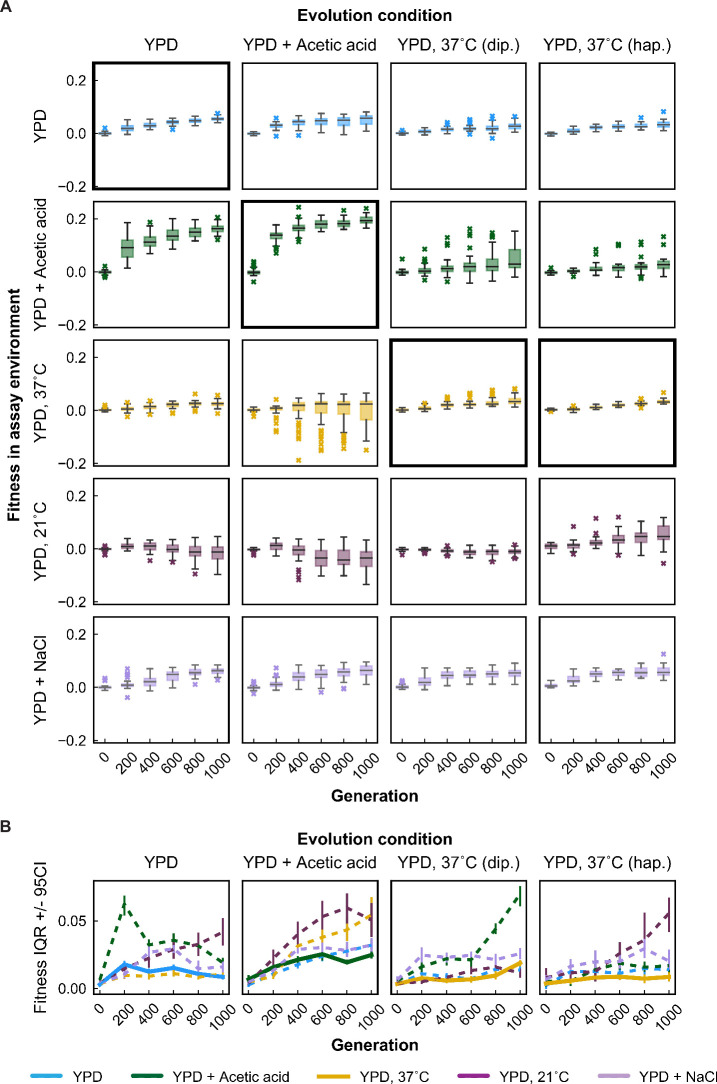
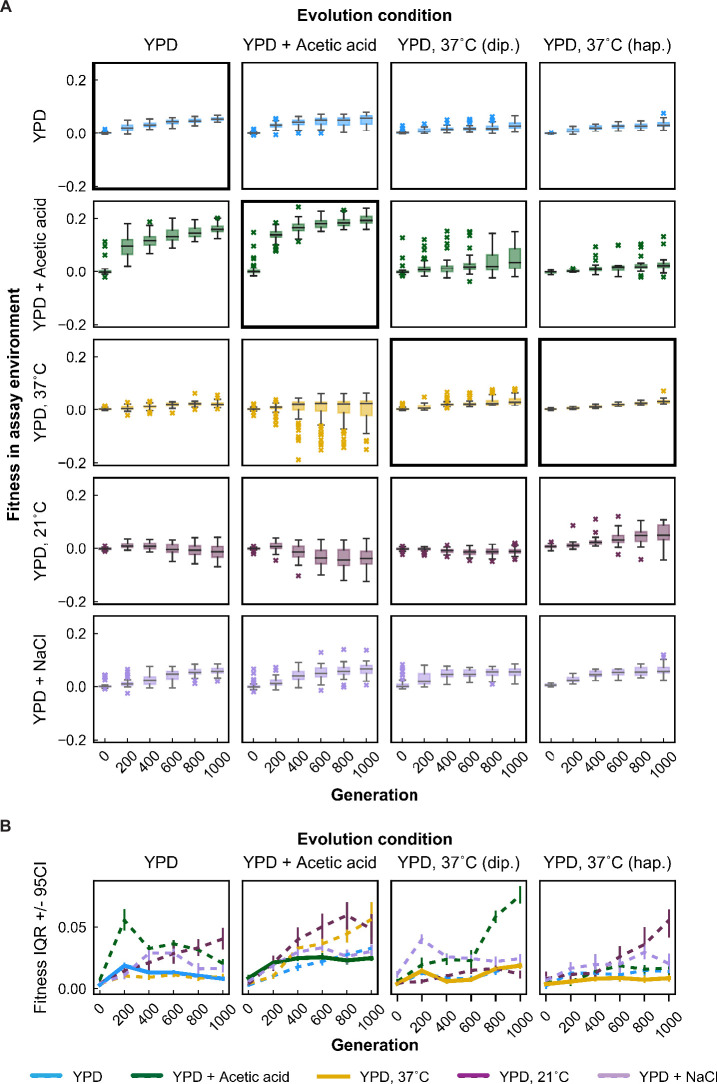
Evolutionary adaptation to a constant environment is driven by the accumulation of mutations which can have a range of unrealized pleiotropic effects in other environments. These pleiotropic consequences of adaptation can influence the emergence of specialists or generalists, and are critical for evolution in temporally or spatially fluctuating environments. While many experiments have examined the pleiotropic effects of adaptation at a snapshot in time, very few have observed the dynamics by which these effects emerge and evolve. Here, we propagated hundreds of diploid and haploid laboratory budding yeast populations in each of three environments, and then assayed their fitness in multiple environments over 1000 generations of evolution. We find that replicate populations evolved in the same condition share common patterns of pleiotropic effects across other environments, which emerge within the first several hundred generations of evolution. However, we also find dynamic and environment-specific variability within these trends: variability in pleiotropic effects tends to increase over time, with the extent of variability depending on the evolution environment. These results suggest shifting and overlapping contributions of chance and contingency to the pleiotropic effects of adaptation, which could influence evolutionary trajectories in complex environments that fluctuate across space and time.
进化对恒定环境的适应是由突变的积累驱动的,这些突变在其他环境中可能具有一系列未实现的多效性效应。适应的这些多效性后果会影响专家或通才的出现,并且对于在时间或空间上波动的环境中的进化至关重要。虽然许多实验已经在时间的一个快照中研究了适应的多效性影响,但很少有观察到这些影响出现和演变的动态。在这里,我们在三个环境中的每一个中繁殖了数百个二倍体和单倍体实验室出芽酵母种群,然后在 1000 多代进化过程中在多个环境中测定了它们的适应性。我们发现,在相同条件下进化的复制种群在其他环境中具有共同的多效性效应模式,这些模式在进化的最初几百代内出现。然而,我们也发现这些趋势中的动态和环境特异性可变性:多效性效应的可变性随着时间的推移而增加,其可变性程度取决于进化环境。这些结果表明,适应的多效性效应的机会和偶然性的转移和重叠贡献,这可能会影响在跨越空间和时间波动的复杂环境中的进化轨迹。