Burke Neurological Institute, White Plains, New York, United States.
Brain and Mind Research Institute, Weill Medical College of Cornell University, New York, United States.
Elife. 2021 Oct 1;10:e72873. doi: 10.7554/eLife.72873.
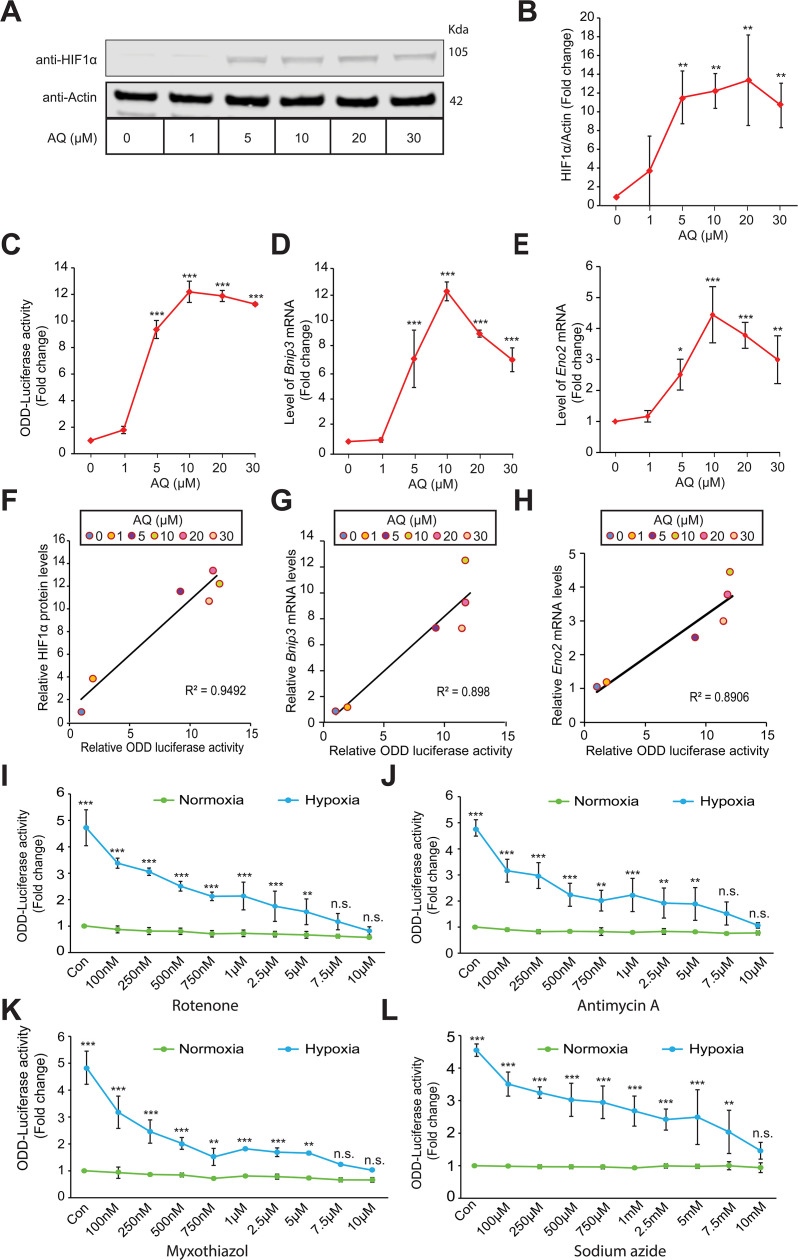
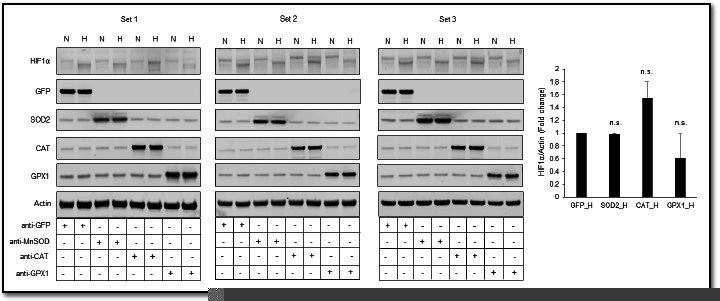
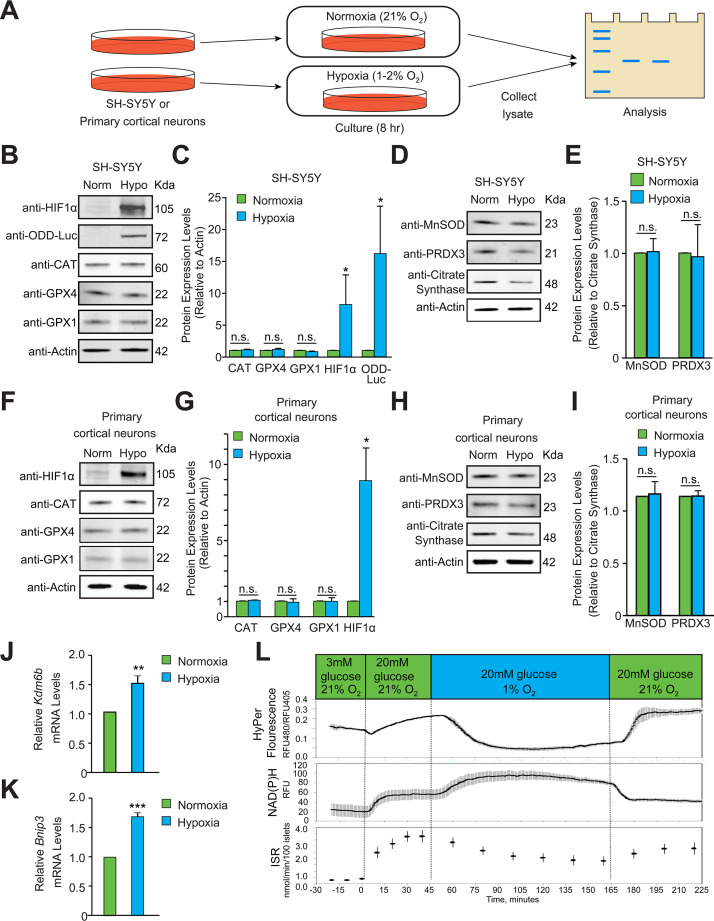
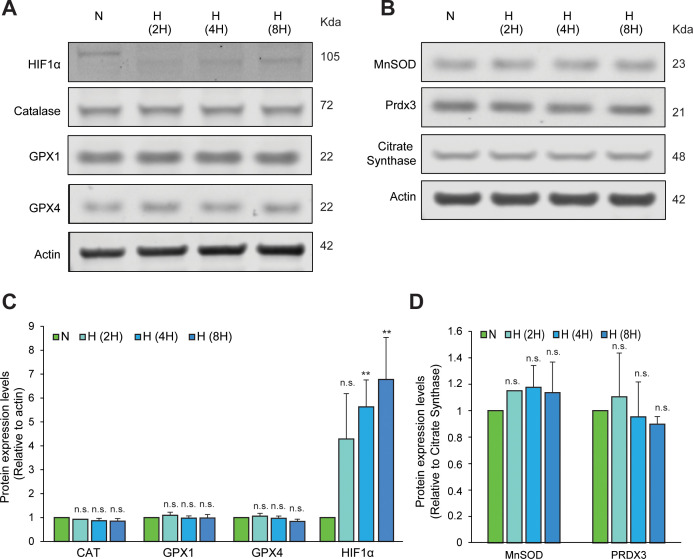
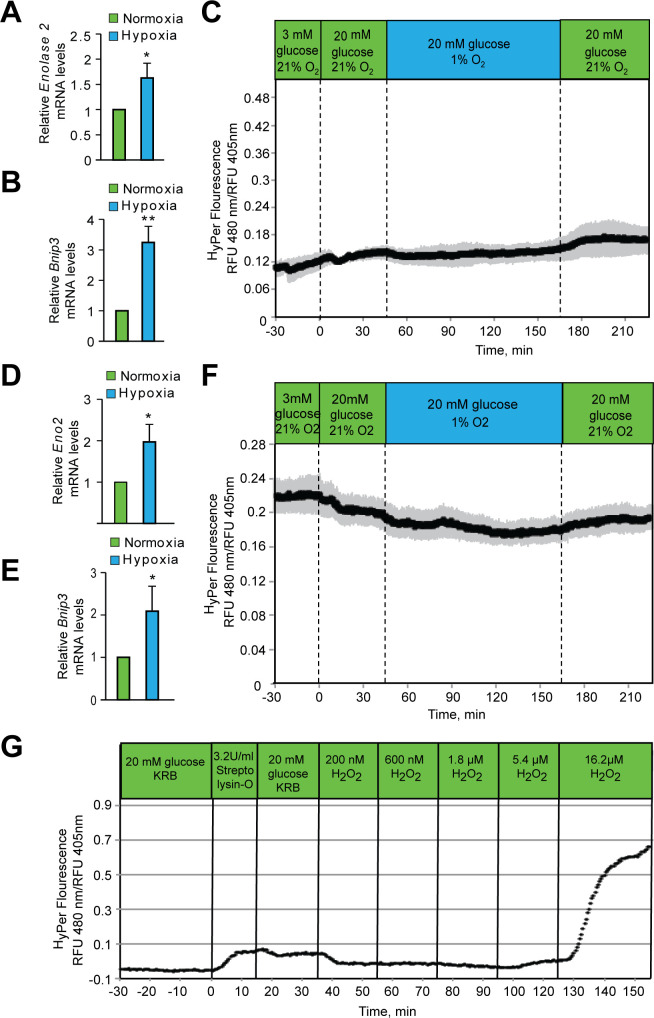
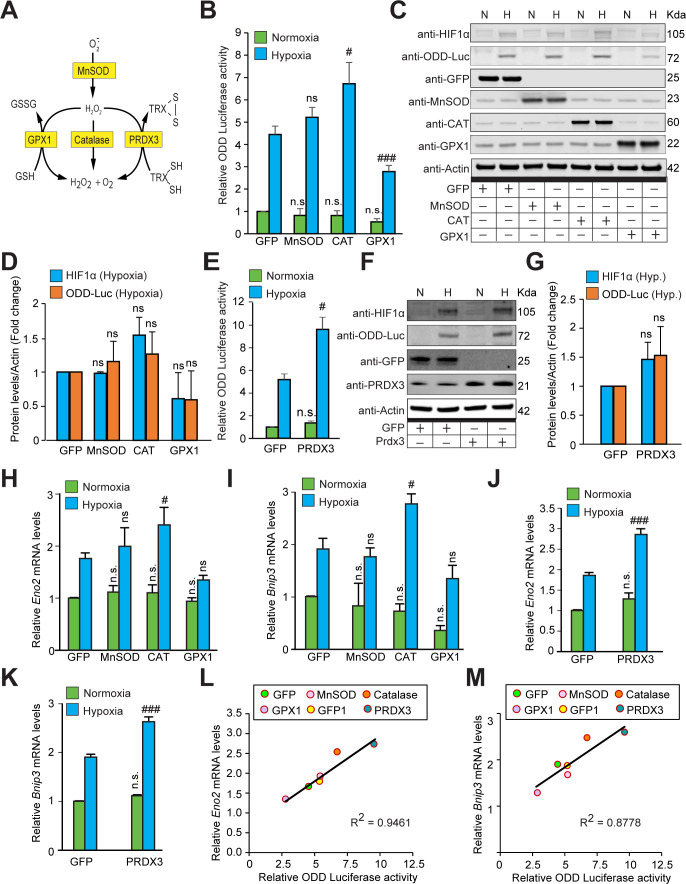
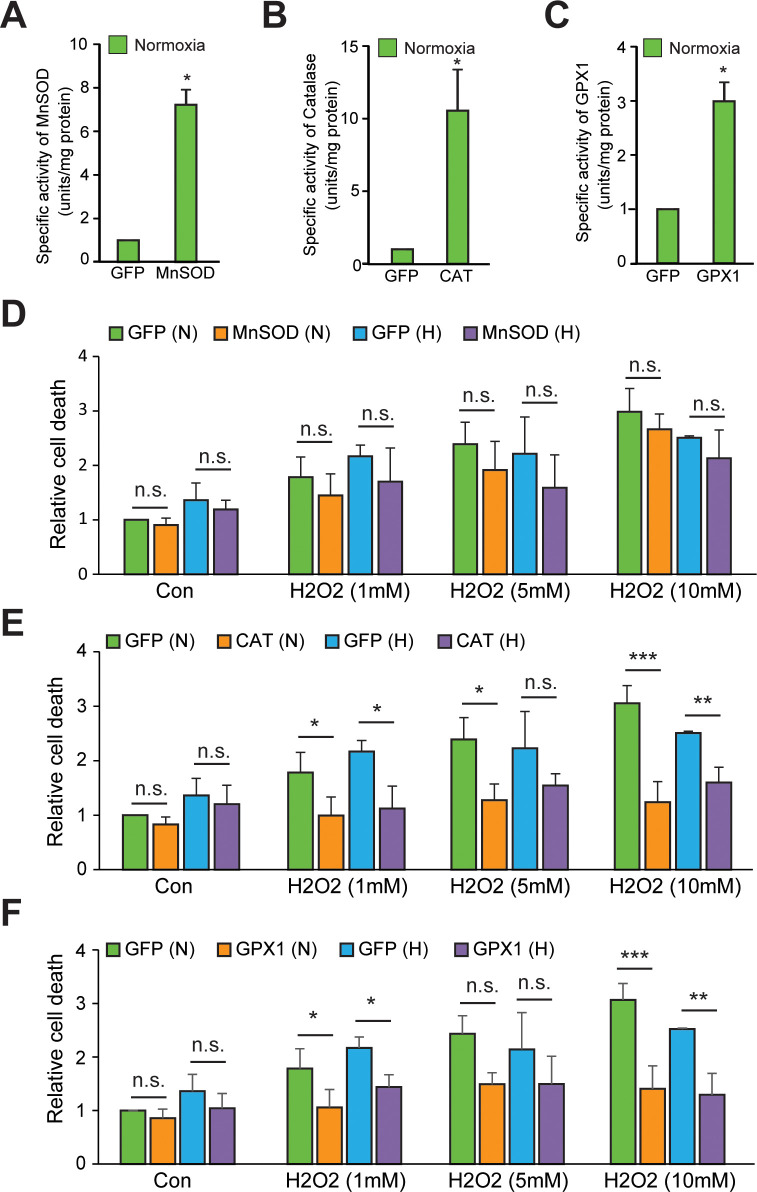
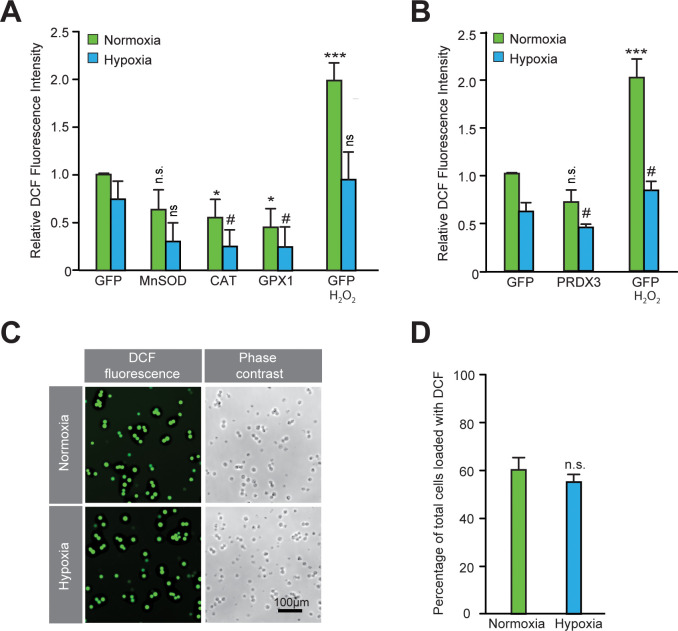
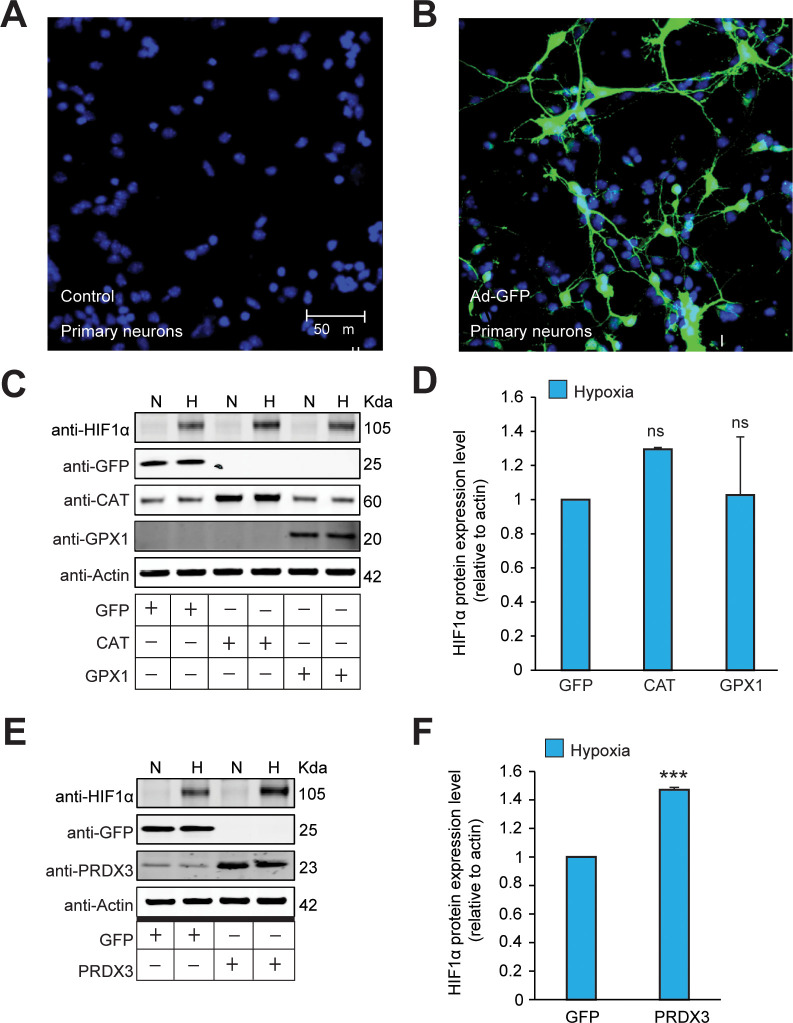
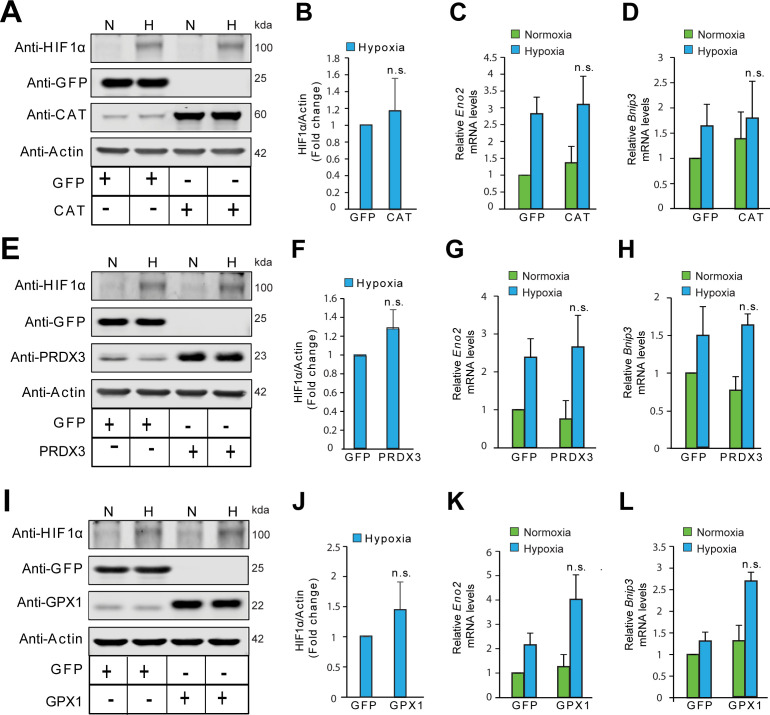
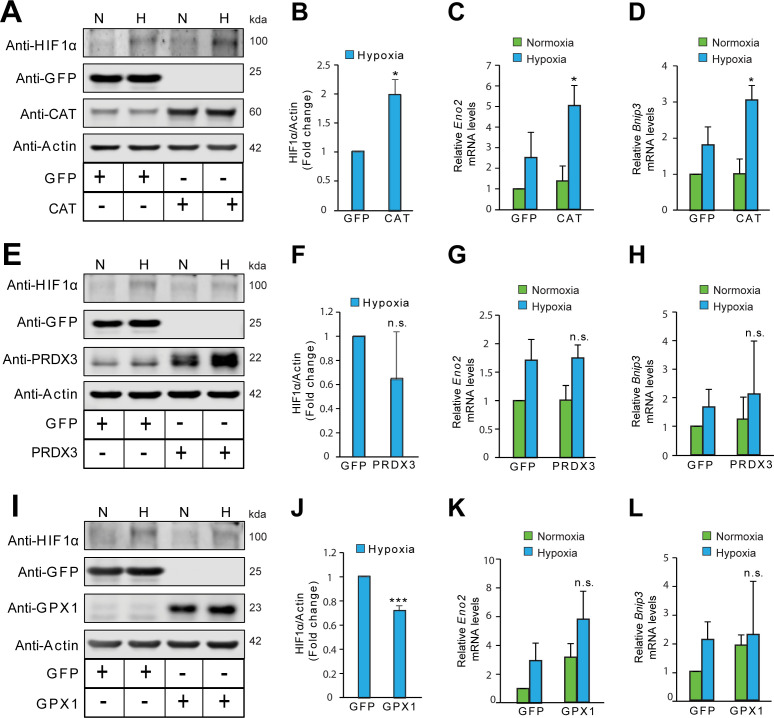
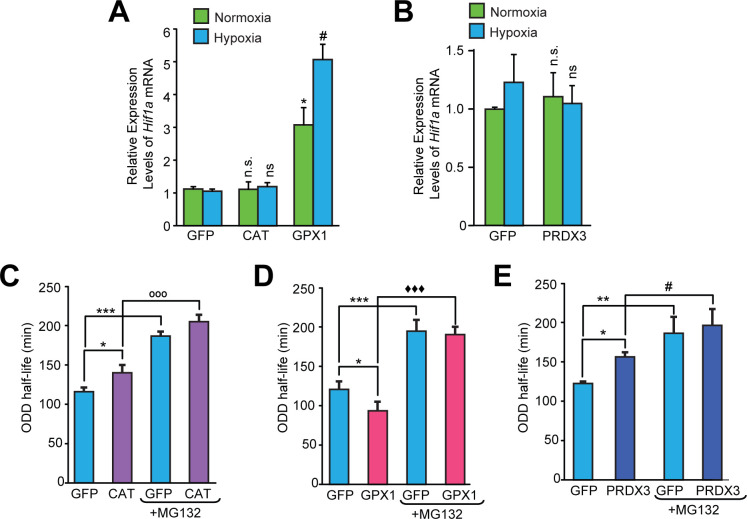
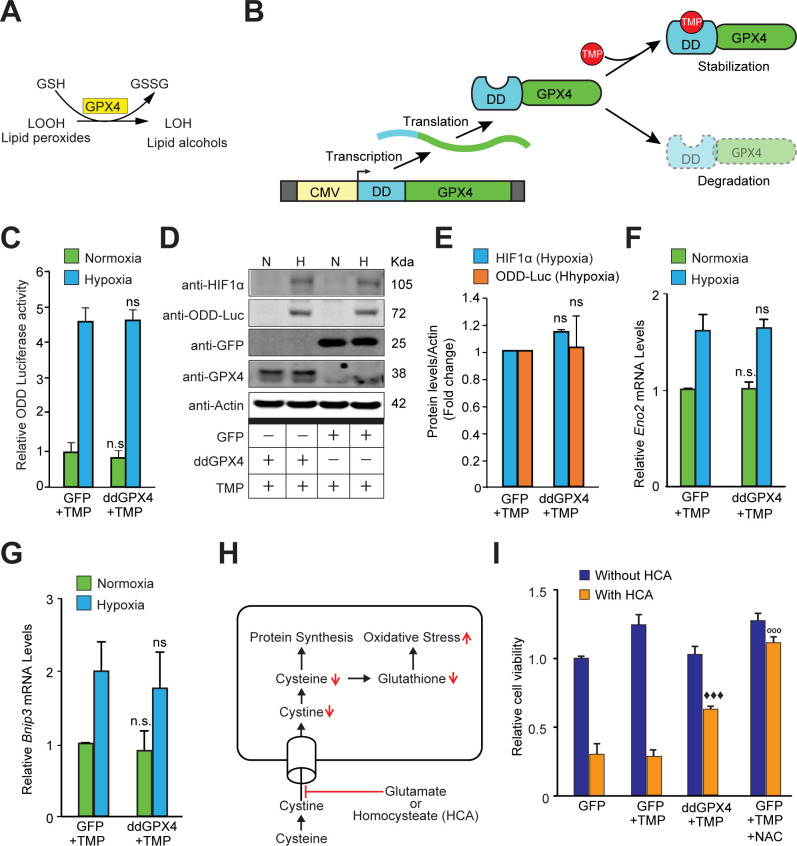
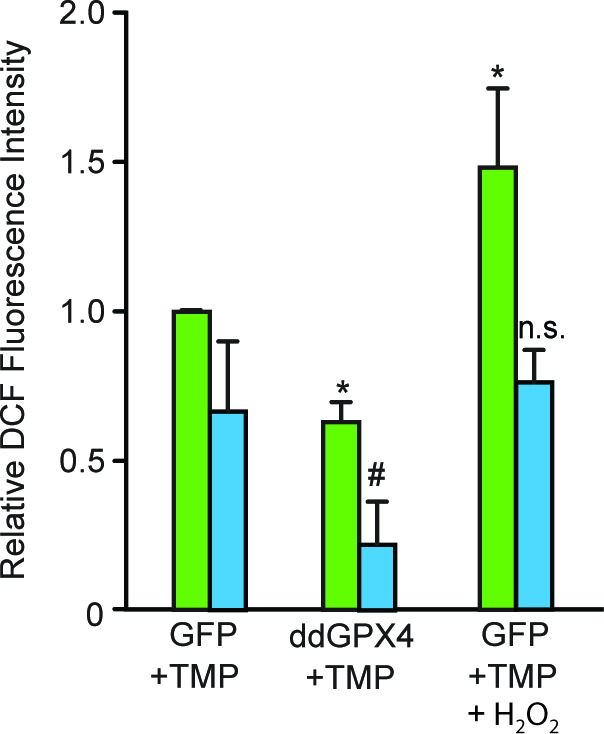
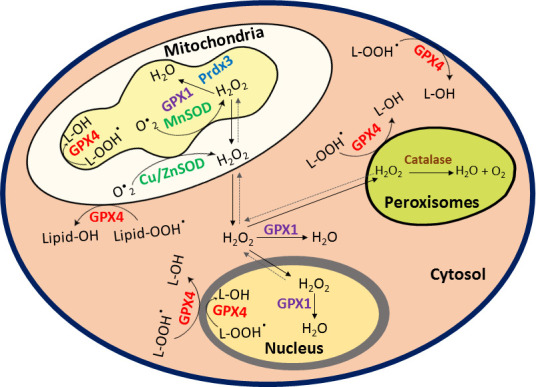
Hypoxic adaptation mediated by HIF transcription factors requires mitochondria, which have been implicated in regulating HIF1α stability in hypoxia by distinct models that involve consuming oxygen or alternatively converting oxygen into the second messenger peroxide. Here, we use a ratiometric, peroxide reporter, HyPer to evaluate the role of peroxide in regulating HIF1α stability. We show that antioxidant enzymes are neither homeostatically induced nor are peroxide levels increased in hypoxia. Additionally, forced expression of diverse antioxidant enzymes, all of which diminish peroxide, had disparate effects on HIF1α protein stability. Moreover, decrease in lipid peroxides by glutathione peroxidase-4 or superoxide by mitochondrial SOD, failed to influence HIF1α protein stability. These data show that mitochondrial, cytosolic or lipid ROS were not necessary for HIF1α stability, and favor a model where mitochondria contribute to hypoxic adaptation as oxygen consumers.
缺氧适应由 HIF 转录因子介导,需要线粒体,不同的模型表明线粒体参与调节缺氧时 HIF1α 的稳定性,这些模型涉及消耗氧气或将氧气转化为第二信使过氧化物。在这里,我们使用比率型过氧化物报告子 HyPer 来评估过氧化物在调节 HIF1α 稳定性中的作用。我们表明,抗氧化酶在缺氧时既没有被同定诱导,过氧化物水平也没有增加。此外,不同抗氧化酶的强制表达,所有这些都能减少过氧化物,但对 HIF1α 蛋白稳定性的影响却大不相同。此外,谷胱甘肽过氧化物酶-4 降低脂质过氧化物或线粒体 SOD 降低超氧化物,都不能影响 HIF1α 蛋白稳定性。这些数据表明,线粒体、细胞质或脂质 ROS 对于 HIF1α 的稳定性不是必需的,并且支持线粒体作为氧气消耗者促进缺氧适应的模型。