Department of Medical Chemistry, Graduate School of Medicine, Kyoto University, Kyoto, Japan.
Graduate School of Biostudies, Kyoto University, Kyoto, Japan.
Elife. 2021 Oct 12;10:e71966. doi: 10.7554/eLife.71966.
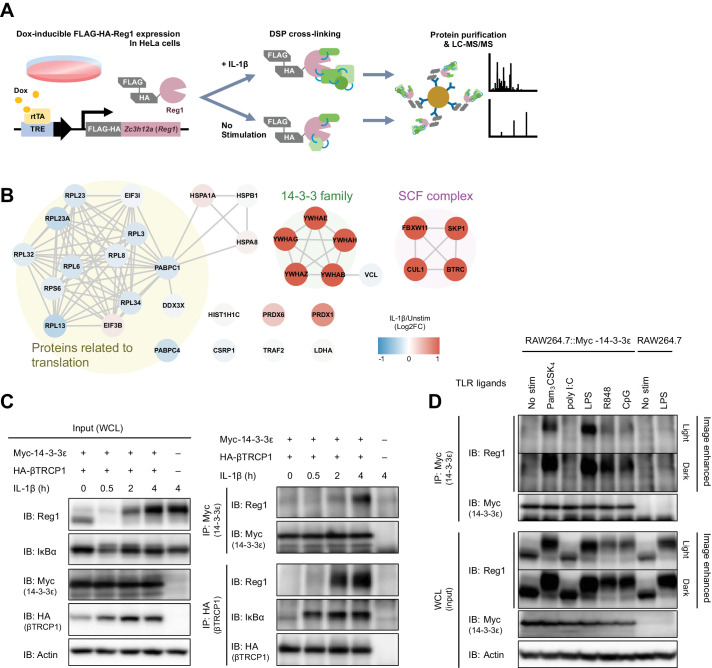
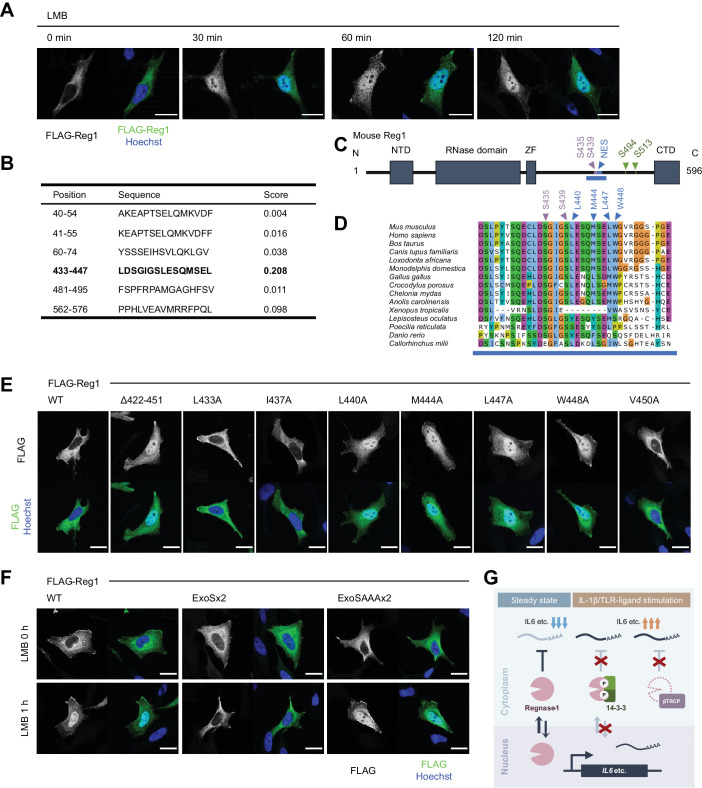
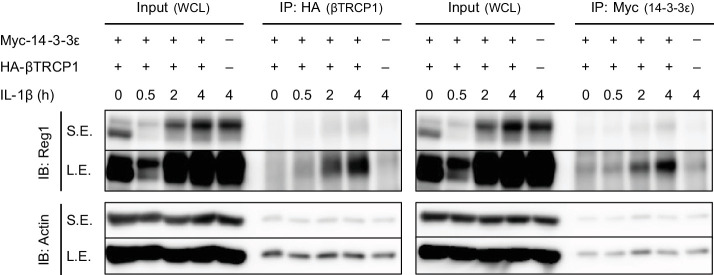
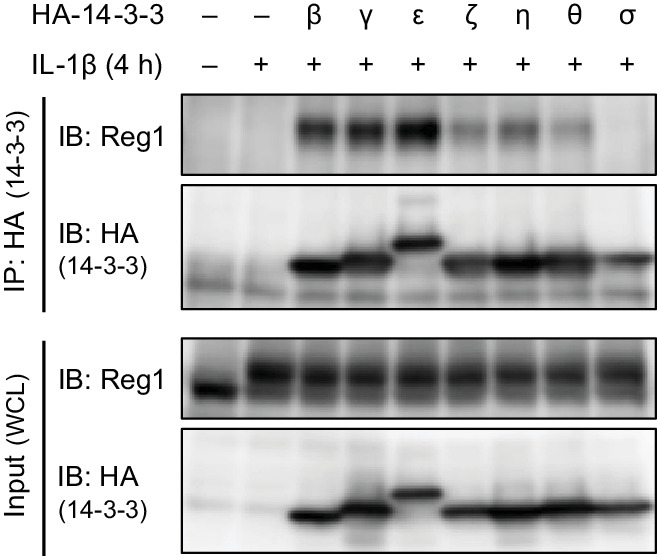
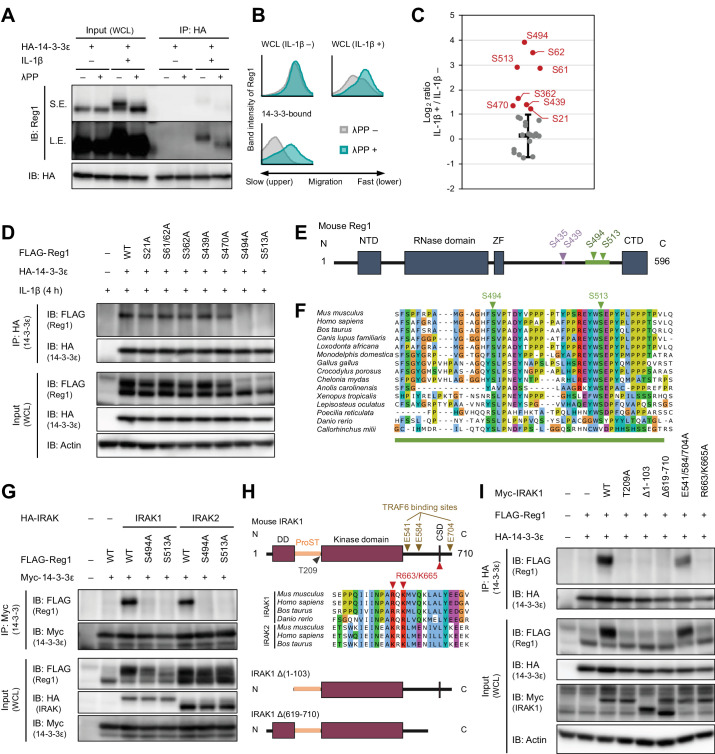
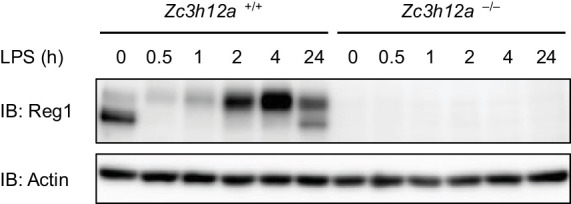
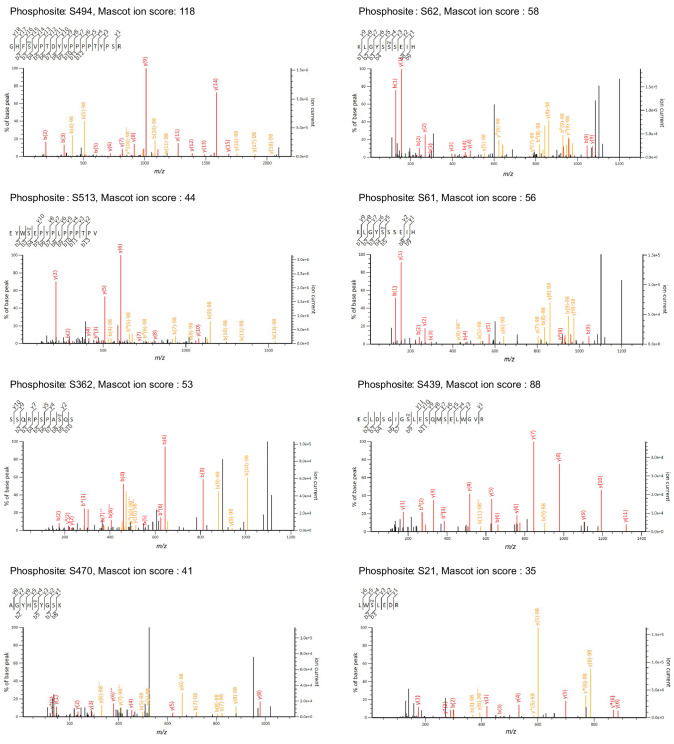
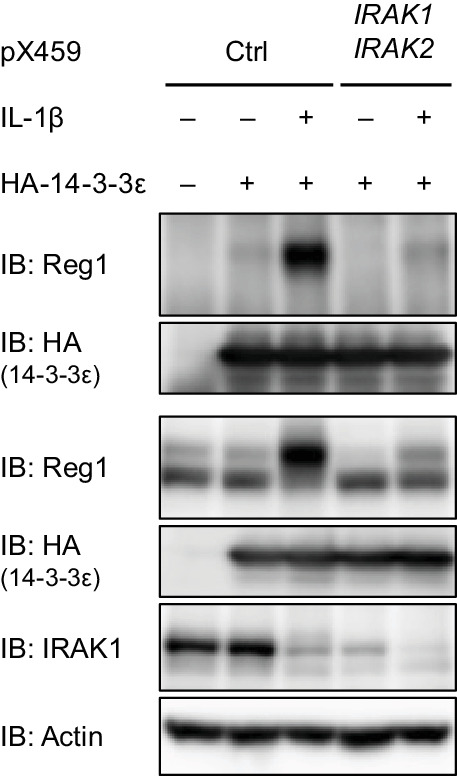
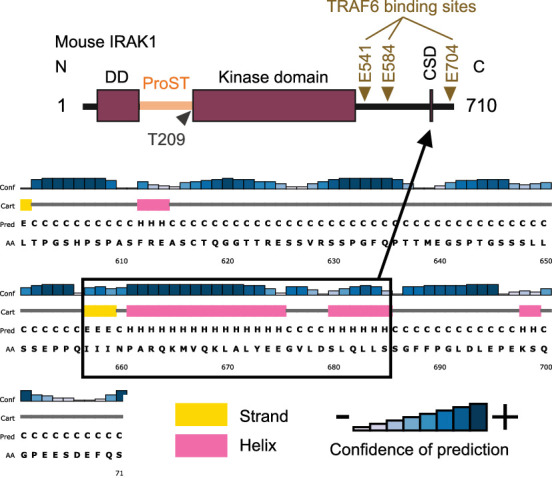
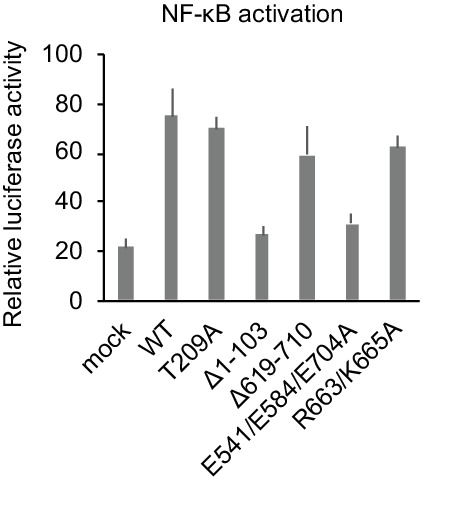
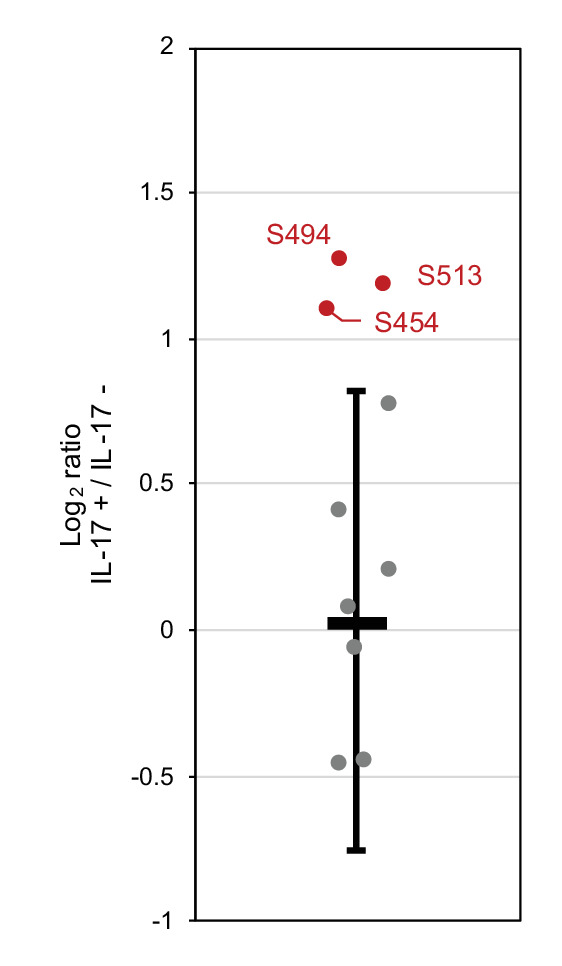
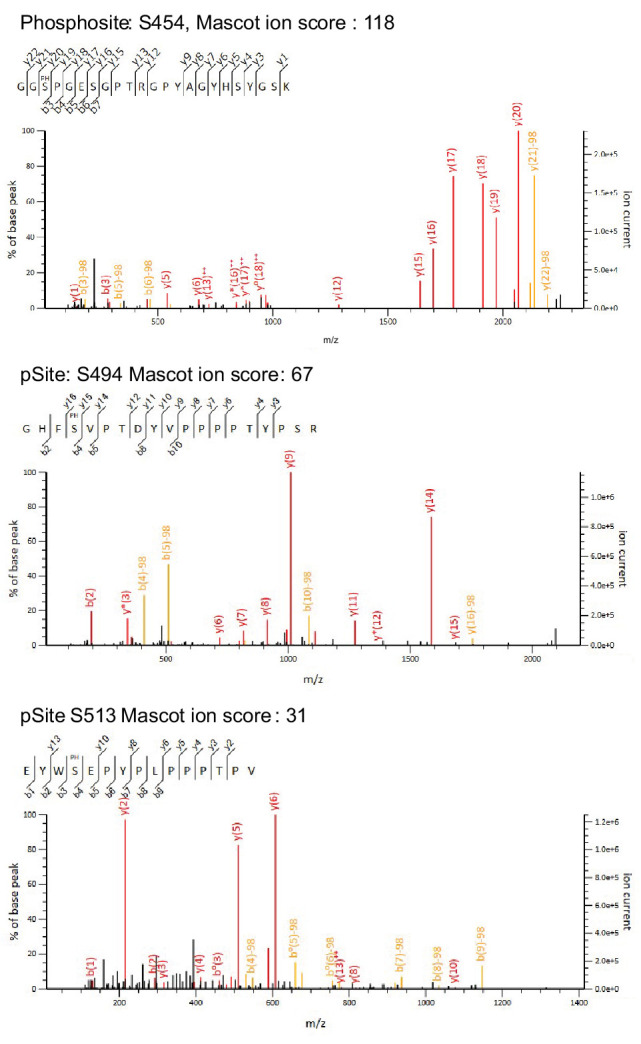
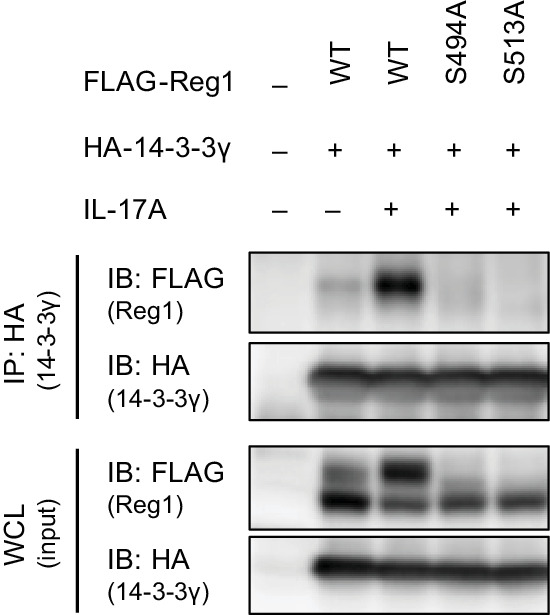
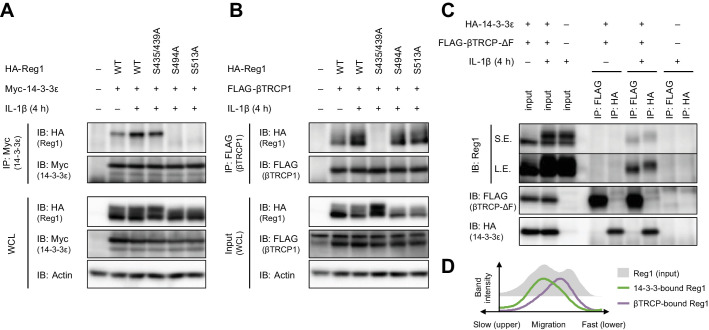
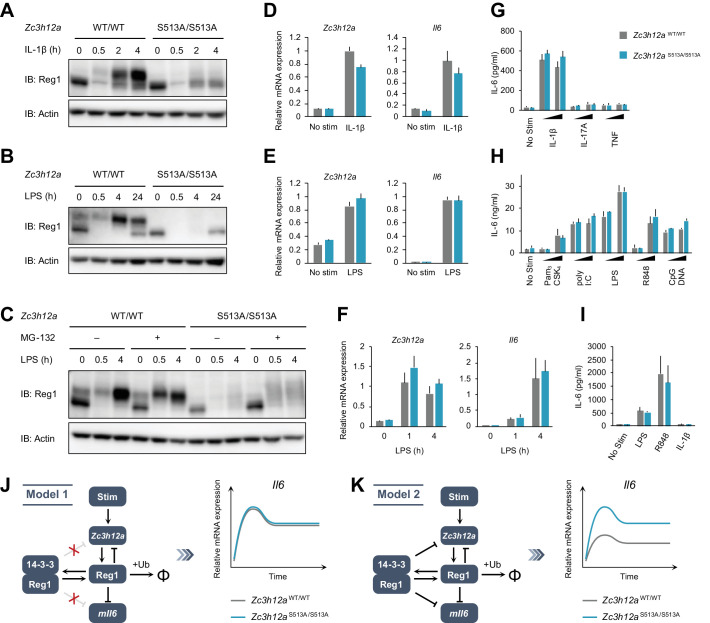
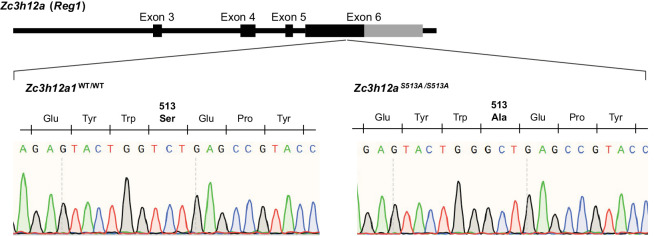
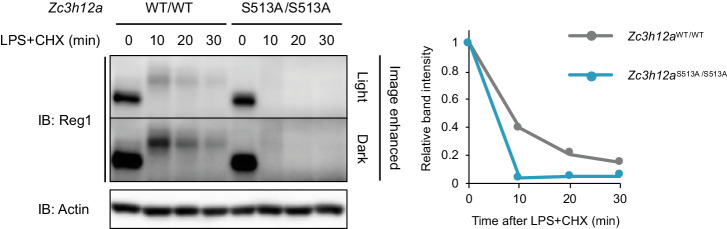
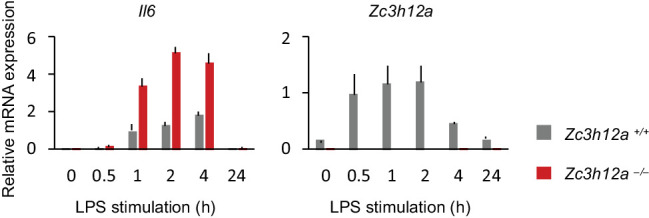
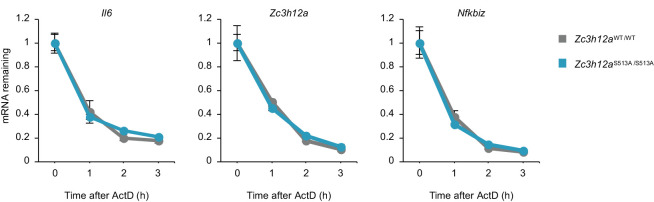
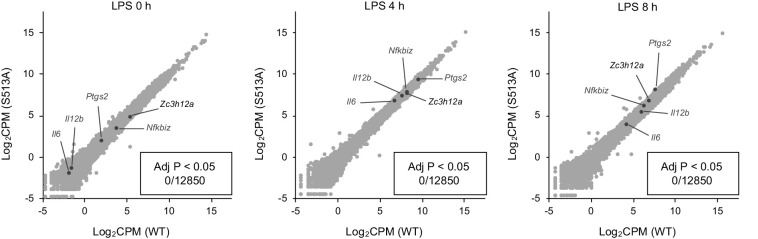
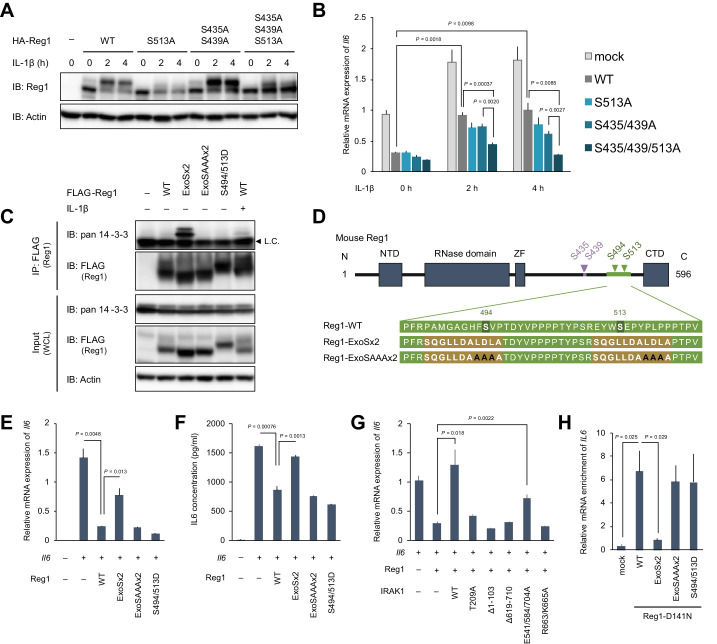
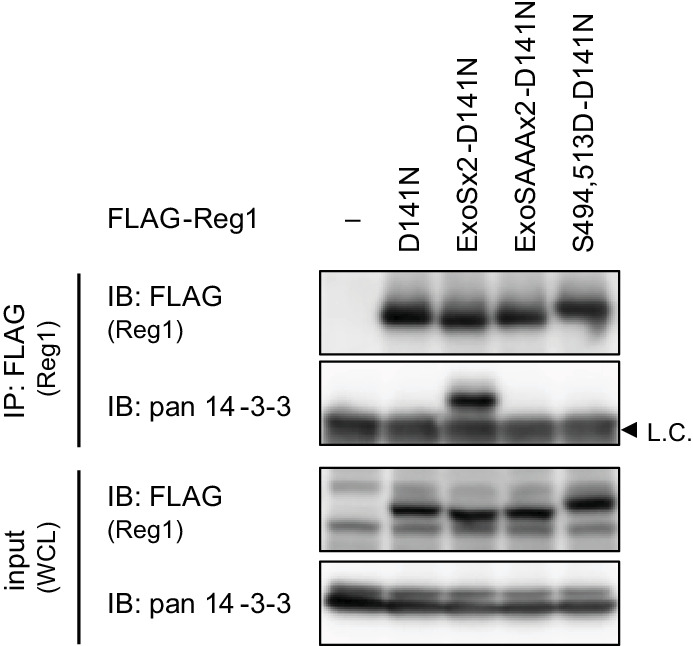
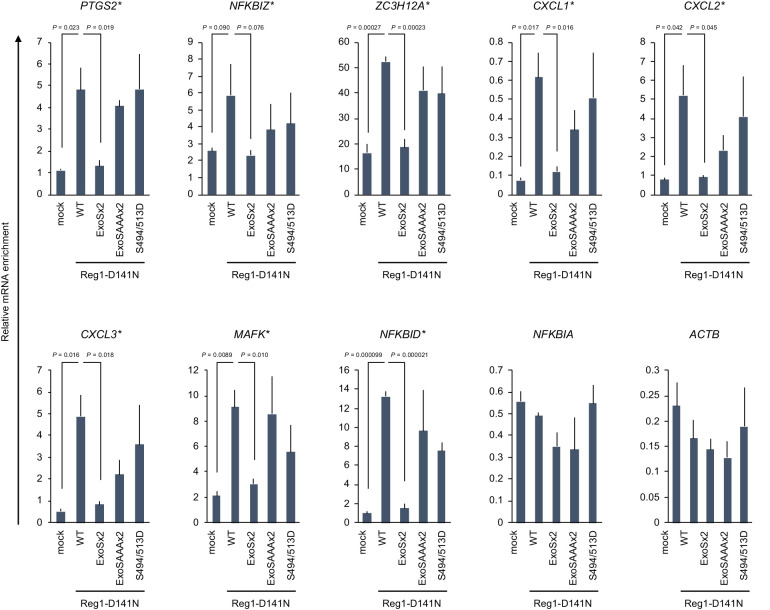
Regnase-1 is an endoribonuclease crucial for controlling inflammation by degrading mRNAs encoding cytokines and inflammatory mediators in mammals. However, it is unclear how Regnase-1-mediated mRNA decay is controlled in interleukin (IL)-1β- or Toll-like receptor (TLR) ligand-stimulated cells. Here, by analyzing the Regnase-1 interactome, we found that IL-1β or TLR stimulus dynamically induced the formation of Regnase-1-β-transducin repeat-containing protein (βTRCP) complex. Importantly, we also uncovered a novel interaction between Regnase-1 and 14-3-3 in both mouse and human cells. In IL-1R/TLR-stimulated cells, the Regnase-1-14-3-3 interaction is mediated by IRAK1 through a previously uncharacterized C-terminal structural domain. Phosphorylation of Regnase-1 at S494 and S513 is critical for Regnase-1-14-3-3 interaction, while a different set of phosphorylation sites of Regnase-1 is known to be required for the recognition by βTRCP and proteasome-mediated degradation. We found that Regnase-1-14-3-3 and Regnase-1-βTRCP interactions are not sequential events. Rather, 14-3-3 protects Regnase-1 from βTRCP-mediated degradation. On the other hand, 14-3-3 abolishes Regnase-1-mediated mRNA decay by inhibiting Regnase-1-mRNA association. In addition, nuclear-cytoplasmic shuttling of Regnase-1 is abrogated by 14-3-3 interaction. Taken together, the results suggest that a novel inflammation-induced interaction of 14-3-3 with Regnase-1 stabilizes inflammatory mRNAs by sequestering Regnase-1 in the cytoplasm to prevent mRNA recognition.
Regnase-1 是一种内切核酸酶,对于哺乳动物通过降解编码细胞因子和炎症介质的 mRNA 来控制炎症至关重要。然而,Regnase-1 介导的 mRNA 降解如何在白细胞介素 (IL)-1β 或 Toll 样受体 (TLR) 配体刺激的细胞中受到控制尚不清楚。在这里,通过分析 Regnase-1 相互作用组,我们发现 IL-1β 或 TLR 刺激动态诱导形成 Regnase-1-β-转导重复蛋白 (βTRCP) 复合物。重要的是,我们还在小鼠和人类细胞中发现了 Regnase-1 和 14-3-3 之间的一种新相互作用。在 IL-1R/TLR 刺激的细胞中,Regnase-1-14-3-3 相互作用是通过 IRAK1 通过一个以前未表征的 C 末端结构域介导的。Regnase-1 的 S494 和 S513 磷酸化对于 Regnase-1-14-3-3 相互作用至关重要,而 Regnase-1 的另一组磷酸化位点已知需要被 βTRCP 和蛋白酶体介导的降解识别。我们发现 Regnase-1-14-3-3 和 Regnase-1-βTRCP 相互作用不是顺序事件。相反,14-3-3 保护 Regnase-1 免受 βTRCP 介导的降解。另一方面,14-3-3 通过抑制 Regnase-1-mRNA 结合来阻止 Regnase-1 介导的 mRNA 降解。此外,Regnase-1 的核质穿梭被 14-3-3 相互作用所阻断。总之,这些结果表明,一种新的炎症诱导的 14-3-3 与 Regnase-1 的相互作用通过将 Regnase-1 隔离在细胞质中来稳定炎症 mRNA,从而防止 mRNA 识别。