Department of Biomedicine, University of Basel, Basel, Switzerland.
Department of Biosystems Science and Engineering, ETH Zurich, Basel, Switzerland.
Life Sci Alliance. 2021 Nov 24;5(2). doi: 10.26508/lsa.202101010. Print 2022 Feb.
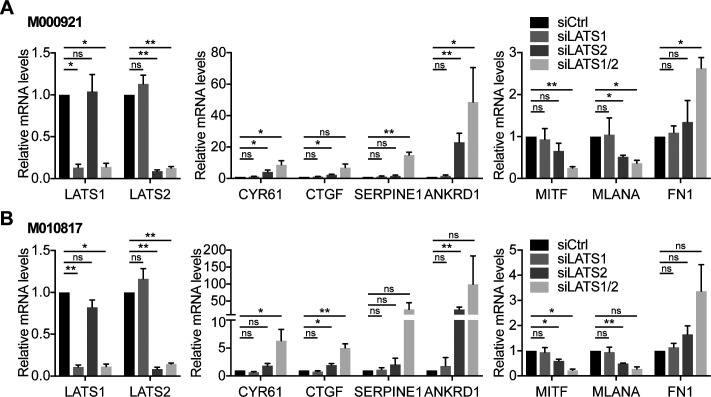
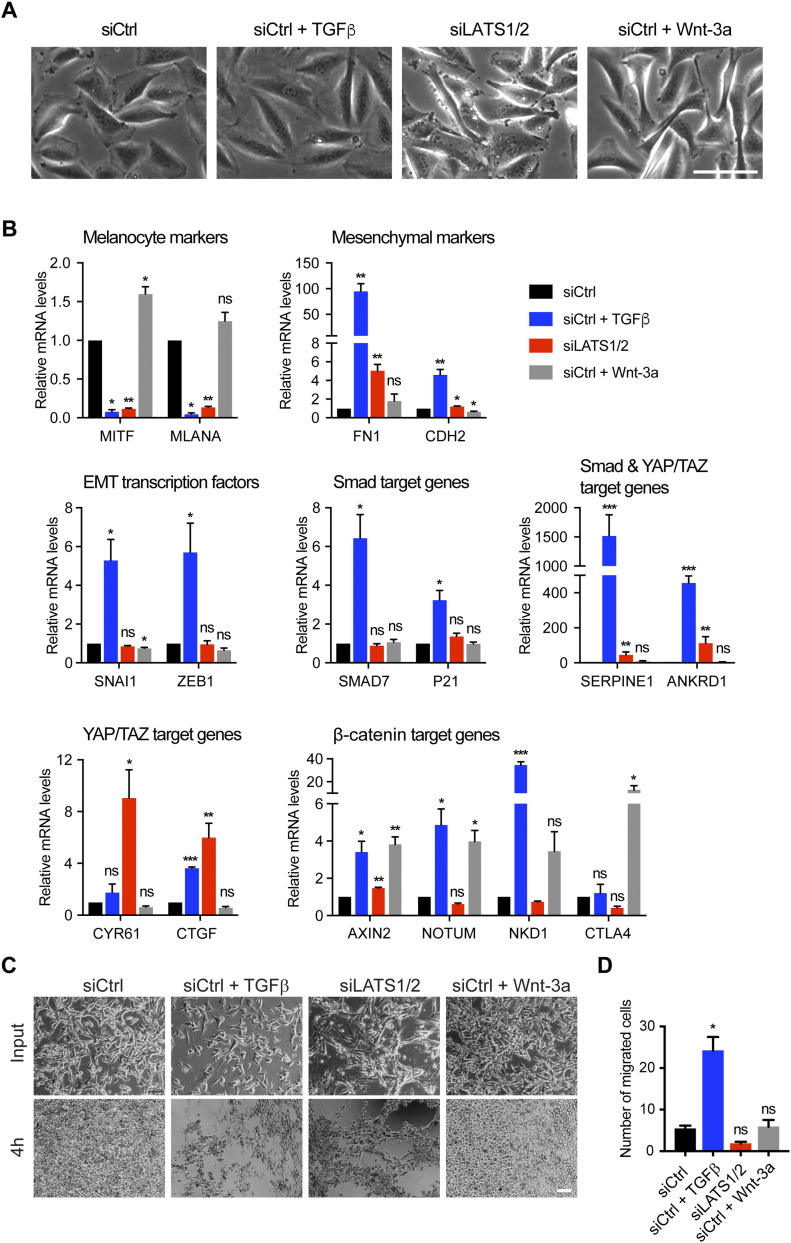
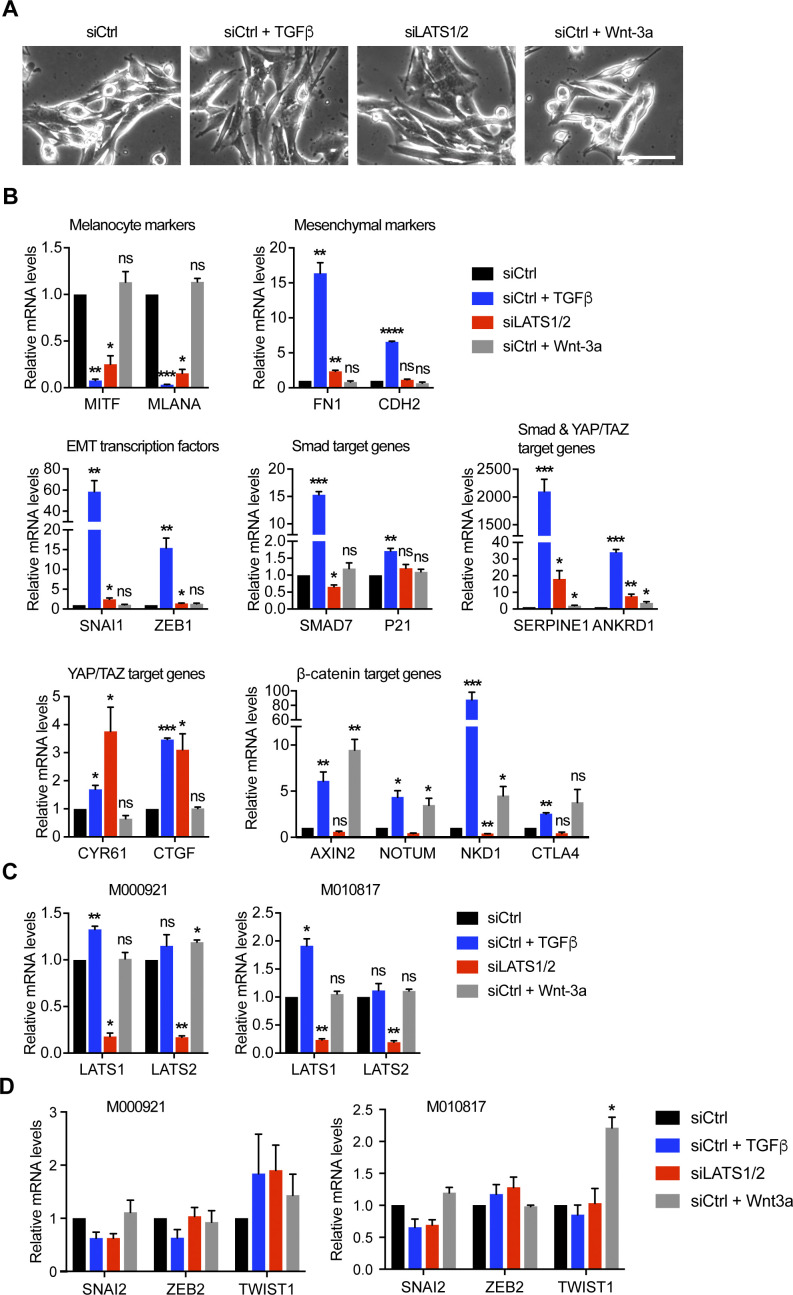
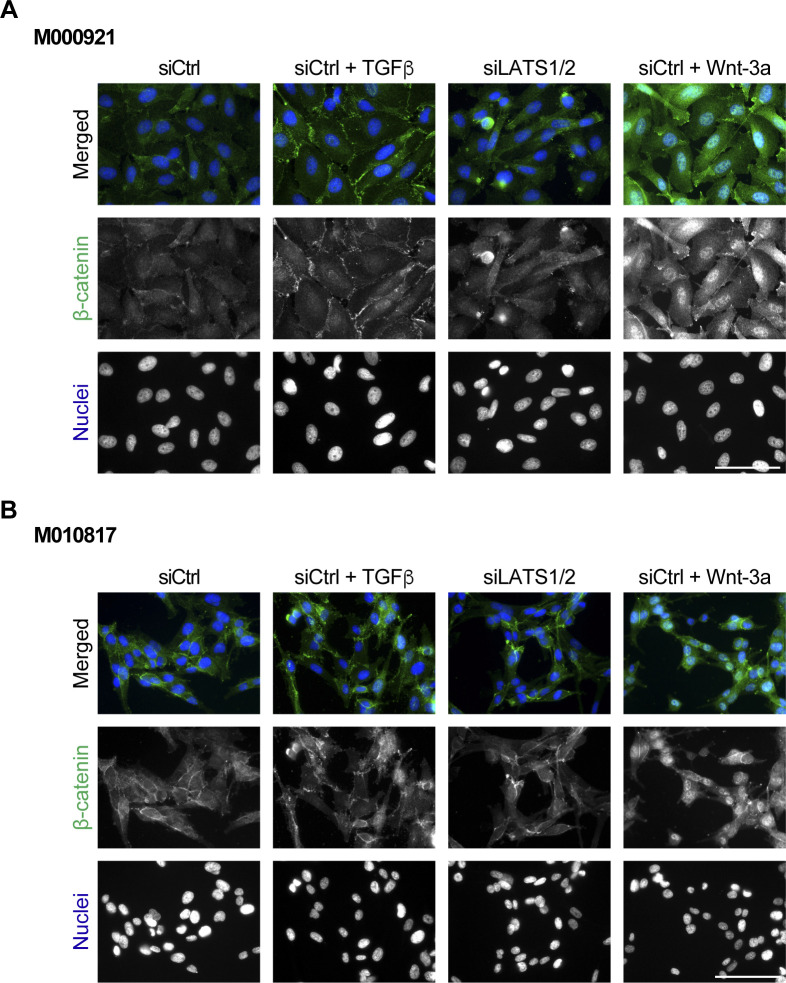
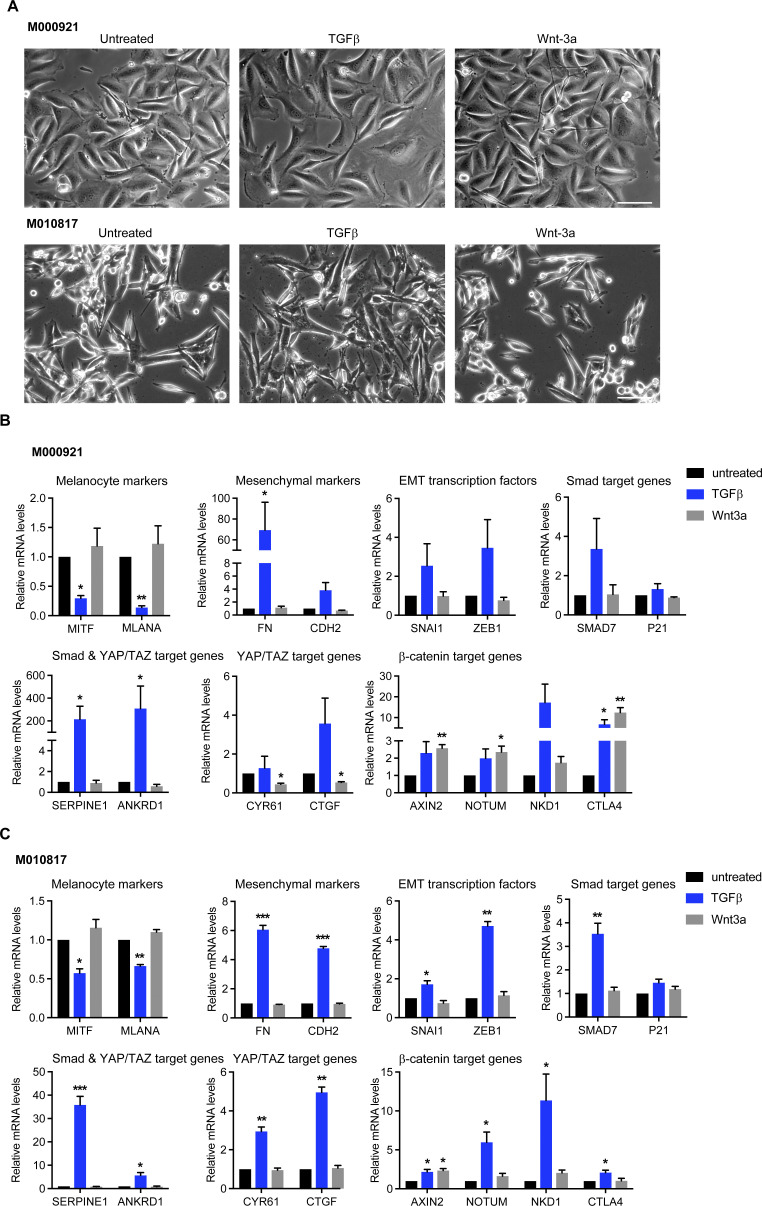
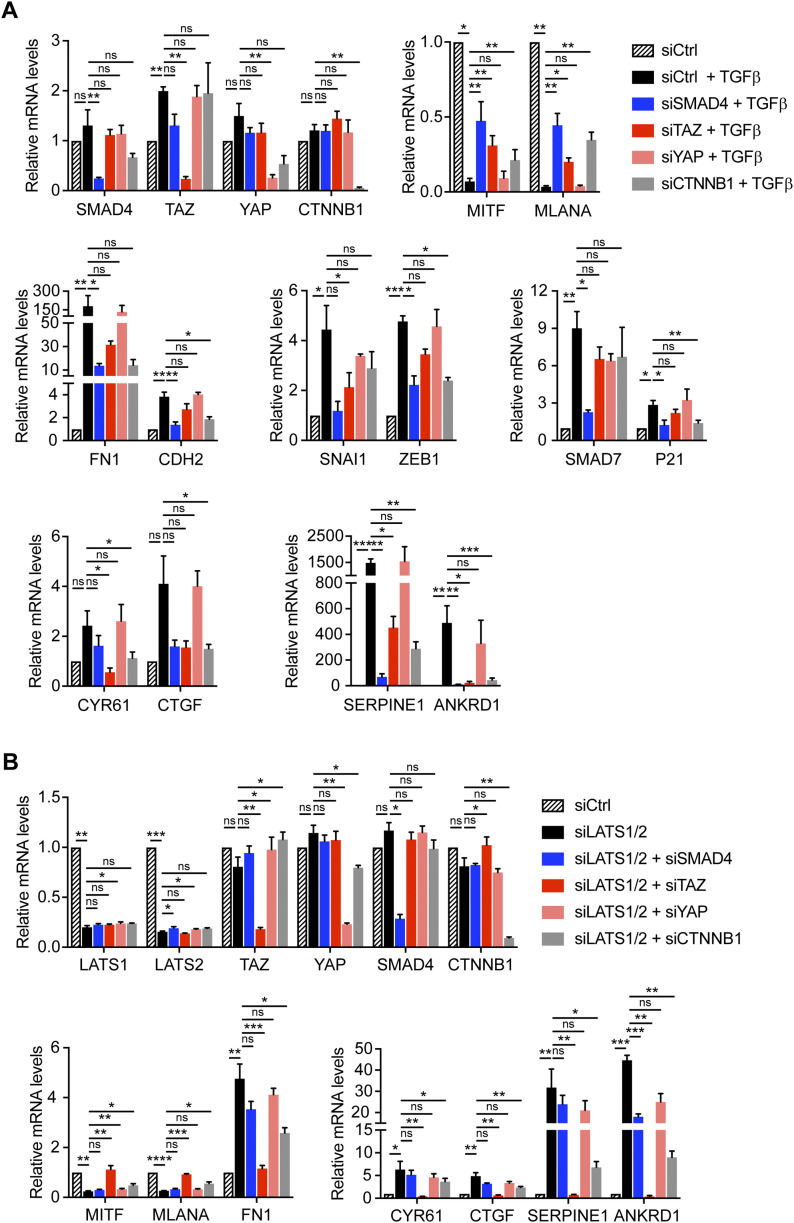
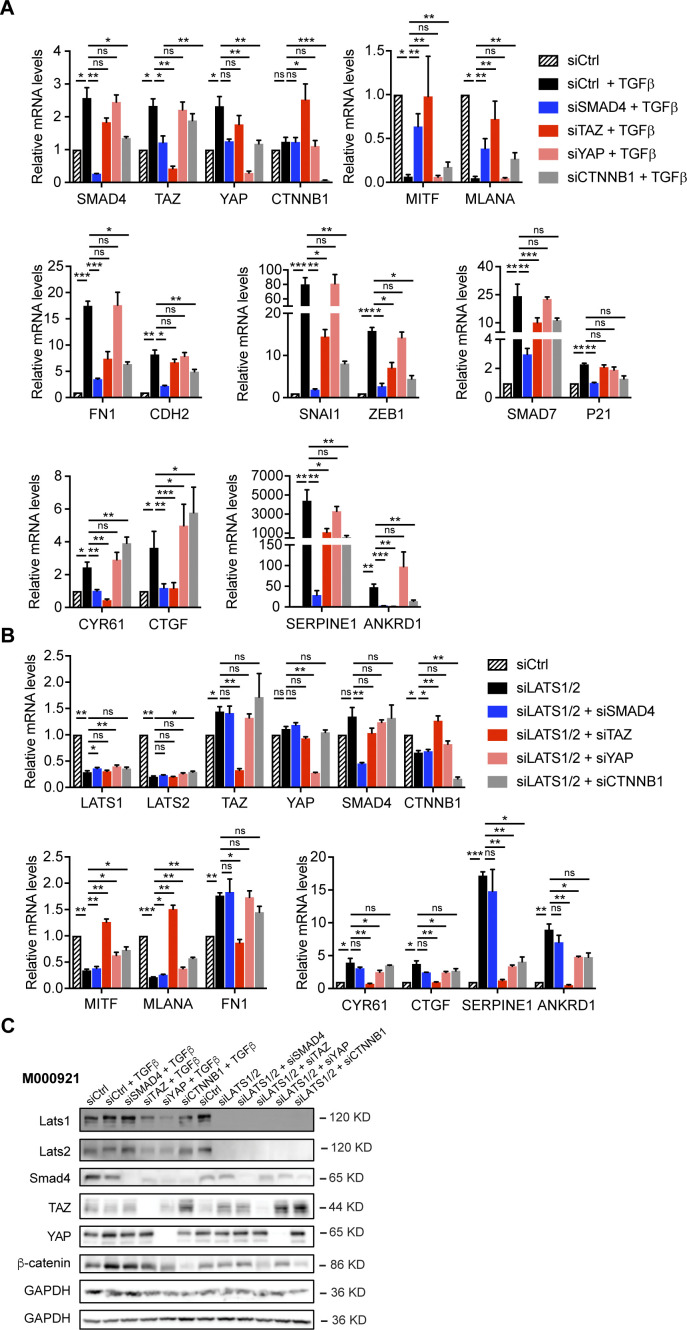
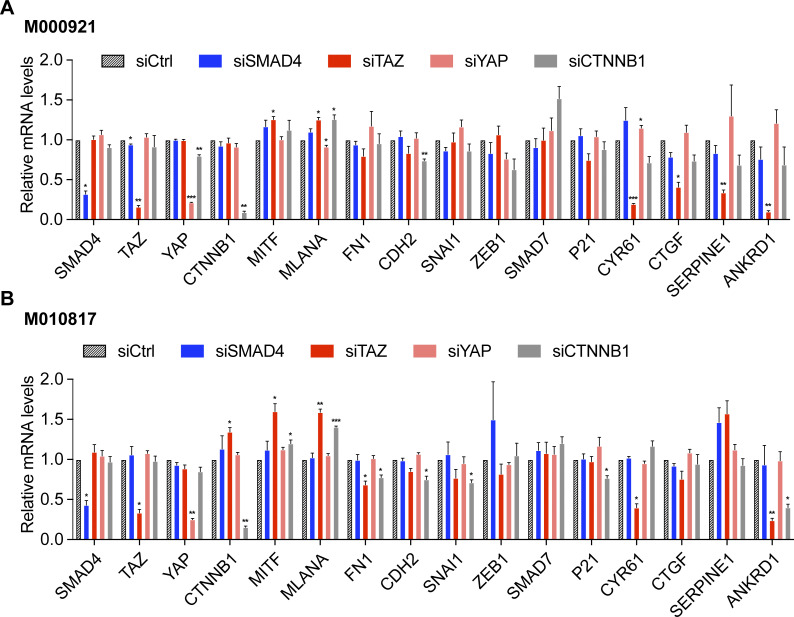
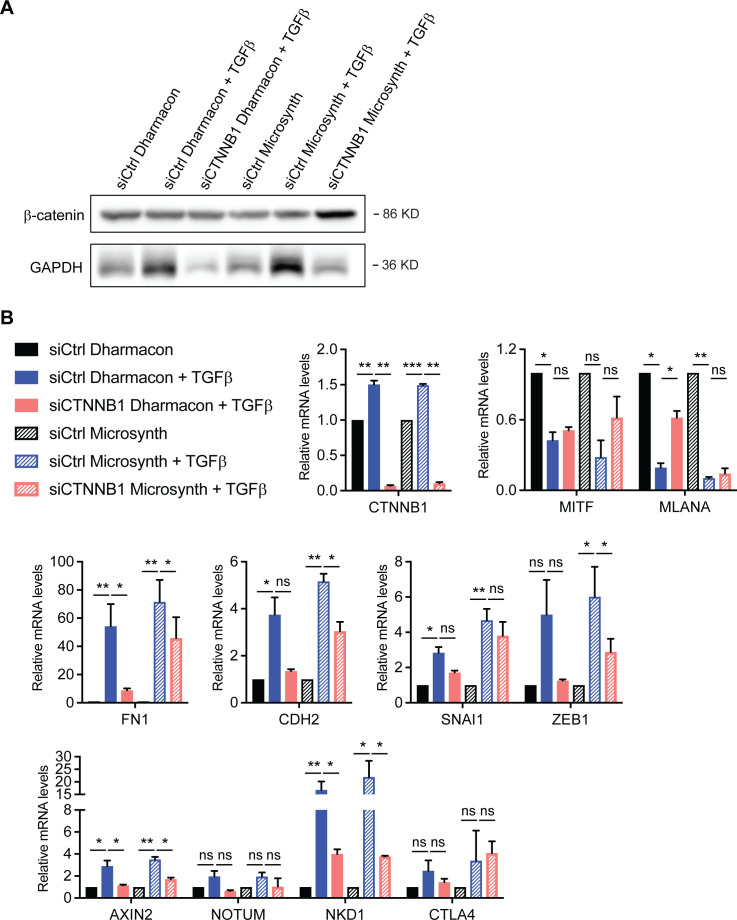
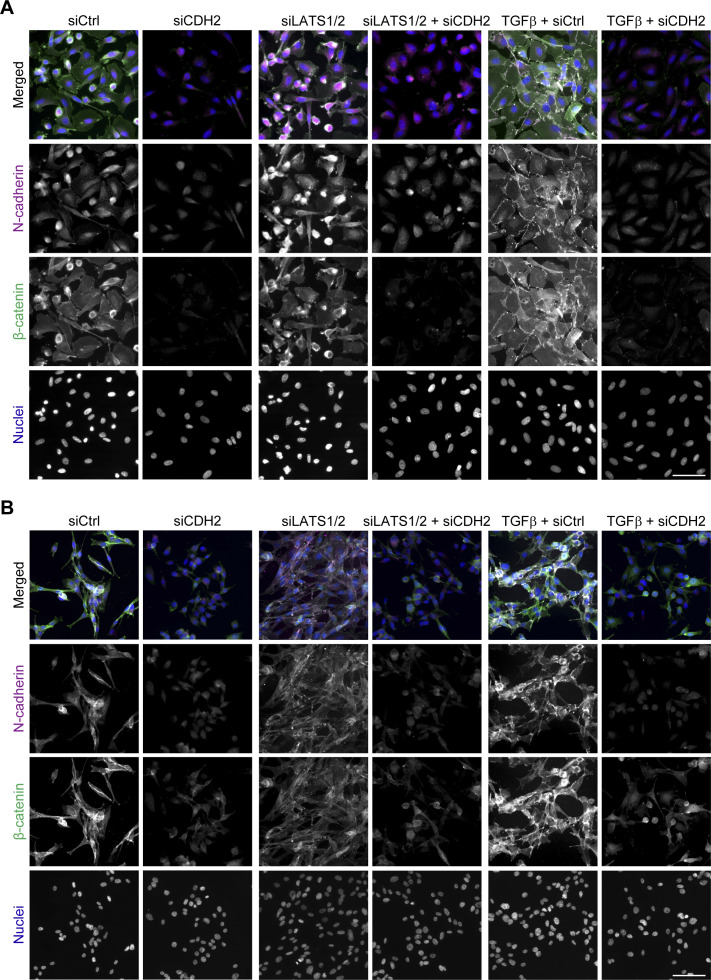
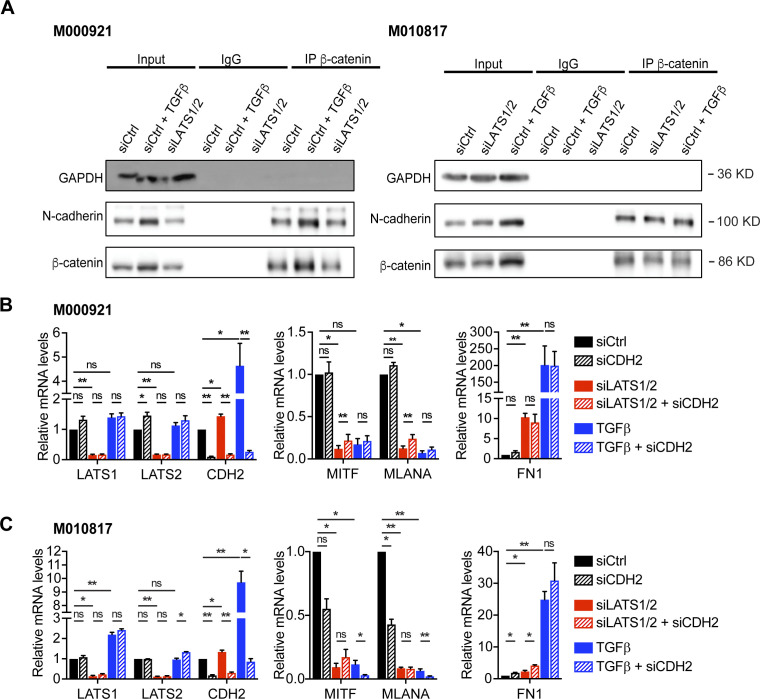
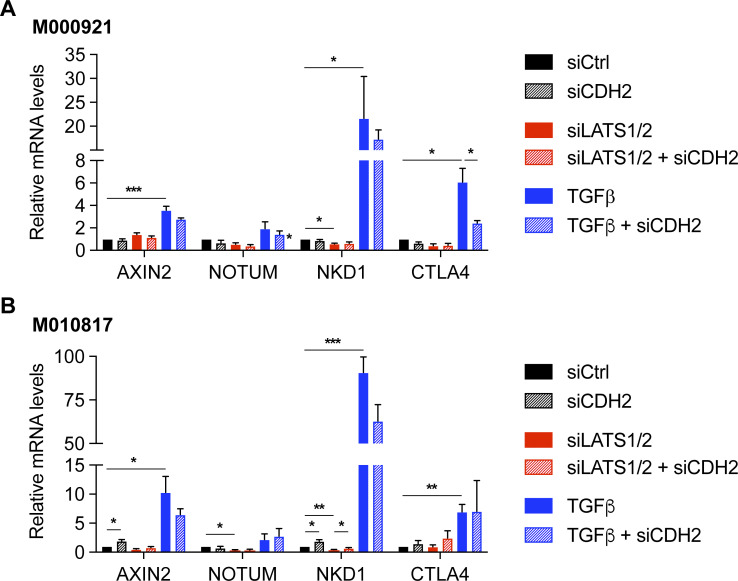
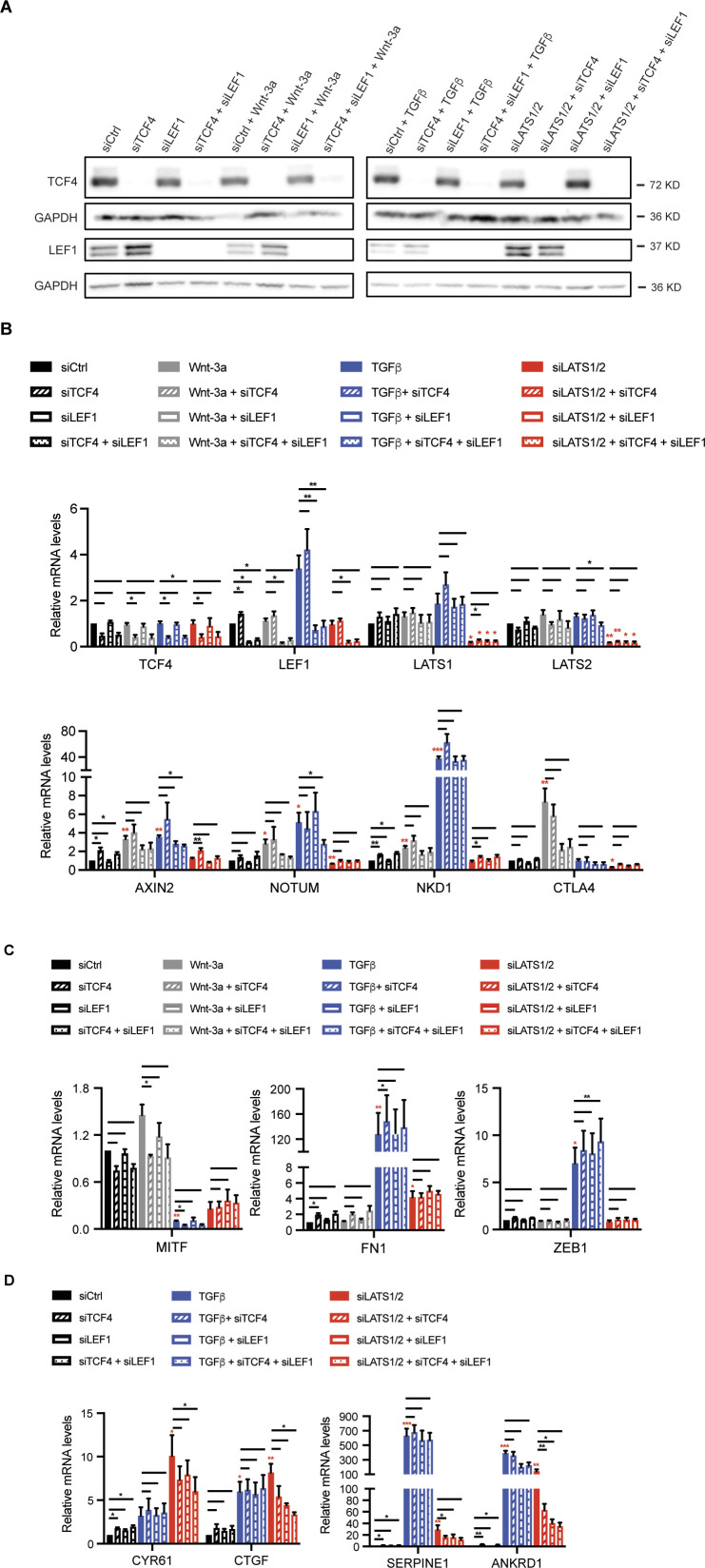
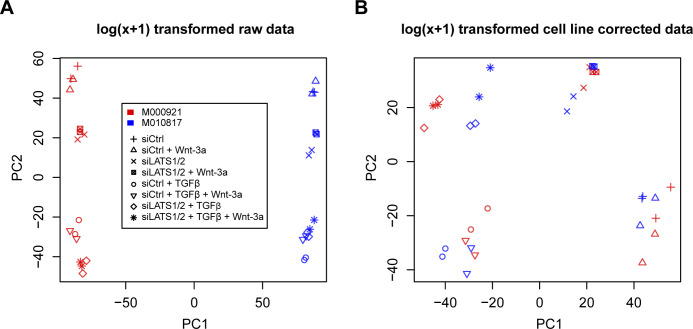
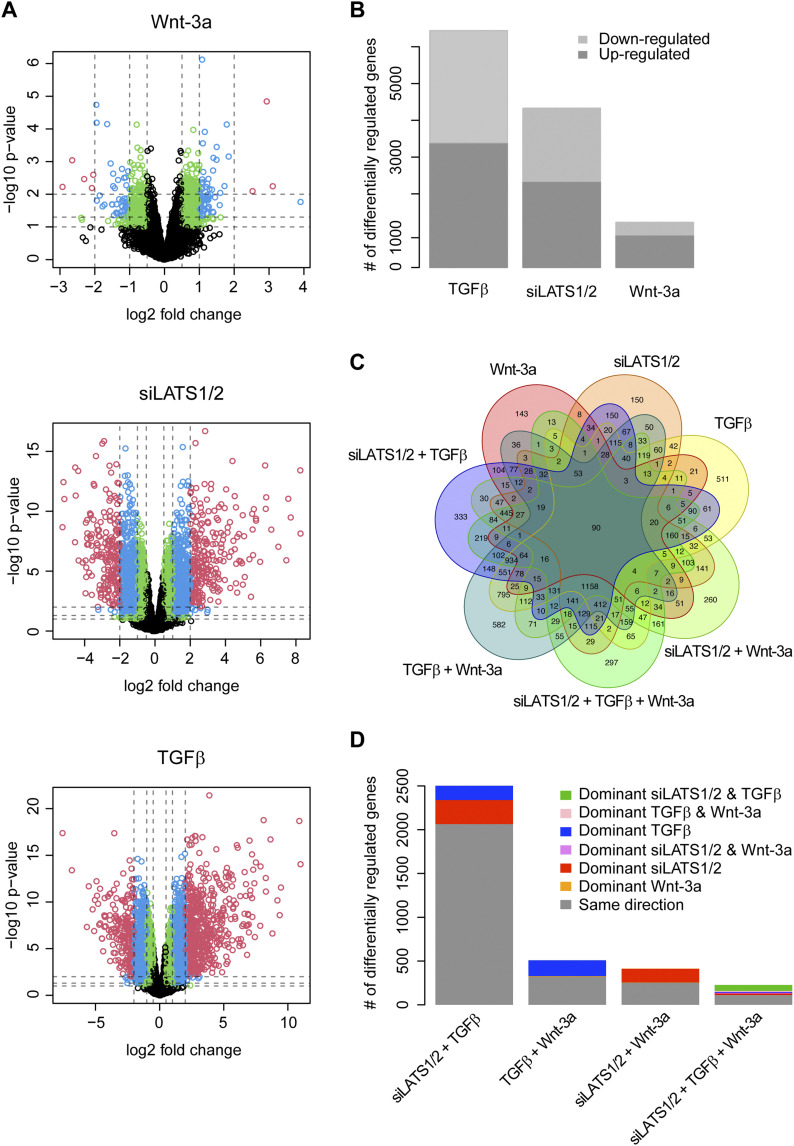
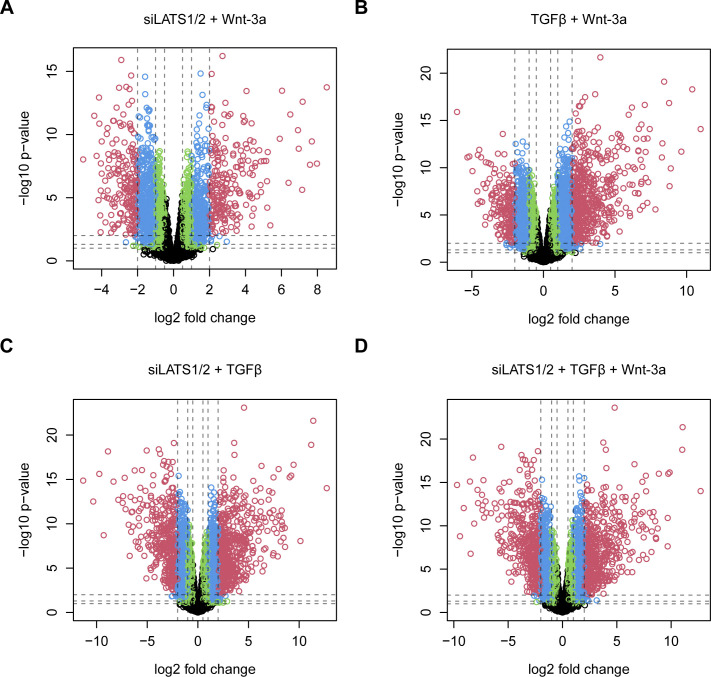
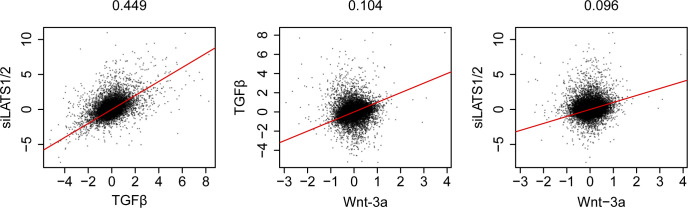
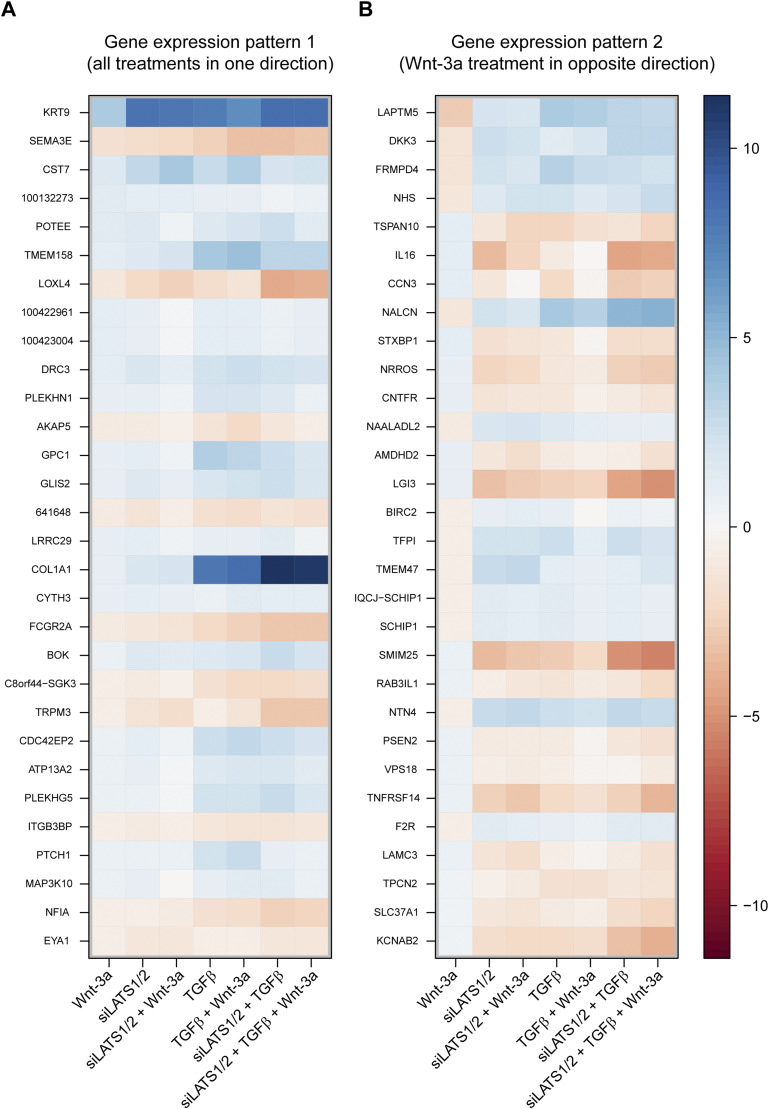
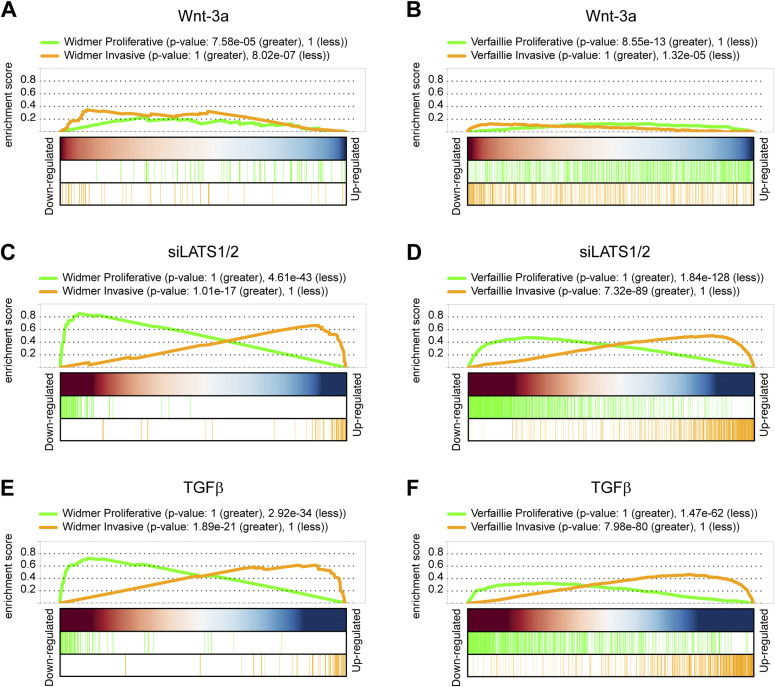
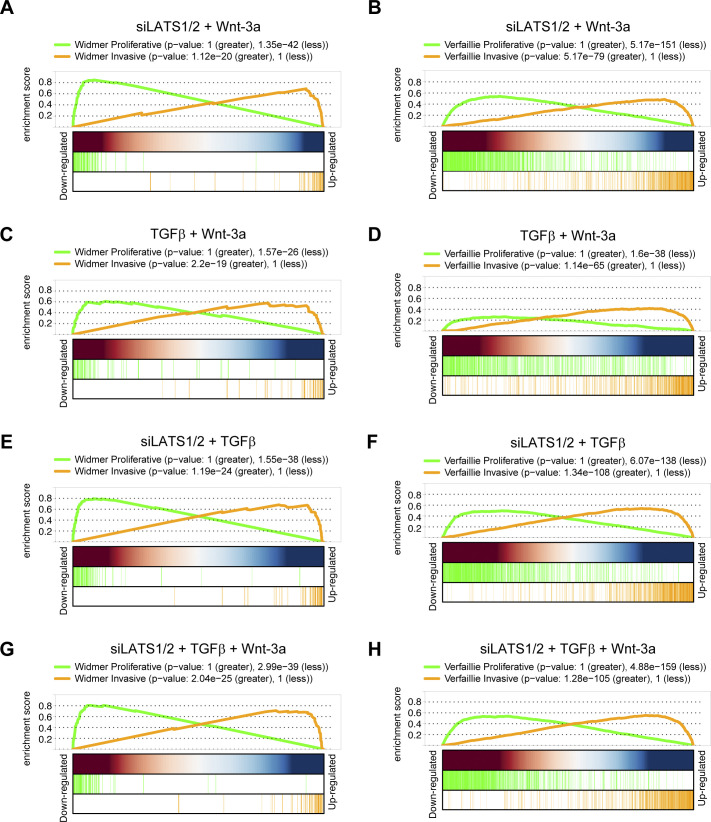
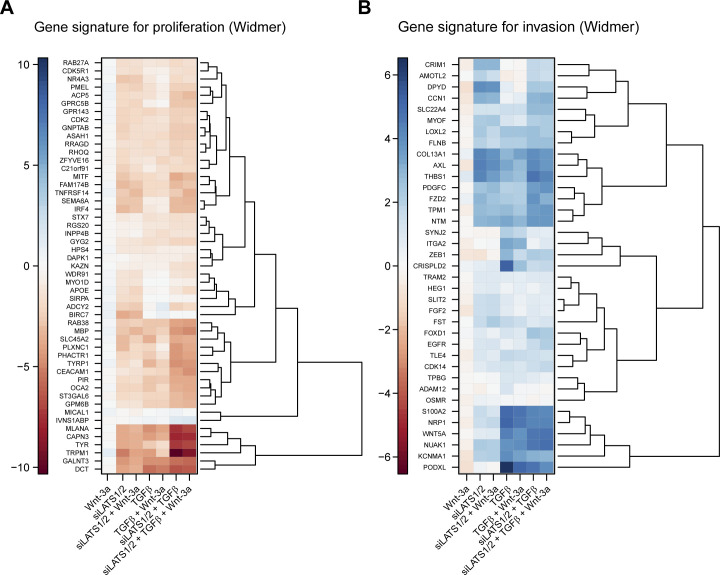
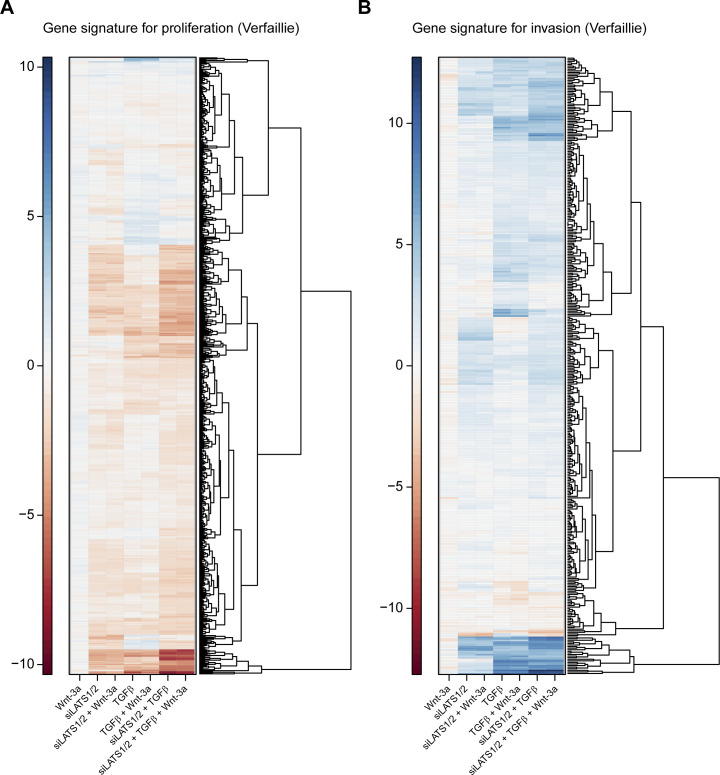
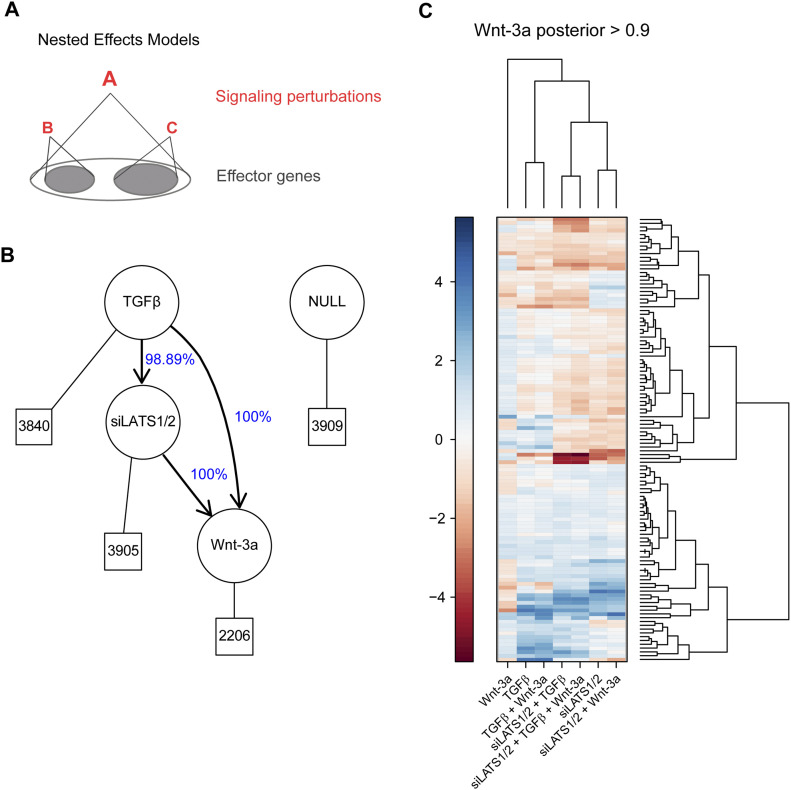
In melanoma, a switch from a proliferative melanocytic to an invasive mesenchymal phenotype is based on dramatic transcriptional reprogramming which involves complex interactions between a variety of signaling pathways and their downstream transcriptional regulators. TGFβ/SMAD, Hippo/YAP/TAZ, and Wnt/β-catenin signaling pathways are major inducers of transcriptional reprogramming and converge at several levels. Here, we report that TGFβ/SMAD, YAP/TAZ, and β-catenin are all required for a proliferative-to-invasive phenotype switch. Loss and gain of function experimentation, global gene expression analysis, and computational nested effects models revealed the hierarchy between these signaling pathways and identified shared target genes. SMAD-mediated transcription at the top of the hierarchy leads to the activation of YAP/TAZ and of β-catenin, with YAP/TAZ governing an essential subprogram of TGFβ-induced phenotype switching. Wnt/β-catenin signaling is situated further downstream and exerts a dual role: it promotes the proliferative, differentiated melanoma cell phenotype and it is essential but not sufficient for SMAD or YAP/TAZ-induced phenotype switching. The results identify epistatic interactions among the signaling pathways underlying melanoma phenotype switching and highlight the priorities in targets for melanoma therapy.
在黑色素瘤中,从增殖性黑素细胞向侵袭性间质表型的转变是基于显著的转录重编程,这涉及多种信号通路及其下游转录调节剂之间的复杂相互作用。TGFβ/SMAD、Hippo/YAP/TAZ 和 Wnt/β-catenin 信号通路是转录重编程的主要诱导因子,并在多个水平上汇聚。在这里,我们报告 TGFβ/SMAD、YAP/TAZ 和 β-catenin 对于增殖到侵袭表型的转变都是必需的。缺失和功能获得实验、全基因表达分析和计算嵌套效应模型揭示了这些信号通路之间的层次关系,并确定了共同的靶基因。位于层次结构顶端的 SMAD 介导的转录导致 YAP/TAZ 和 β-catenin 的激活,YAP/TAZ 调控 TGFβ 诱导的表型转换的一个基本亚程序。Wnt/β-catenin 信号位于下游,发挥双重作用:它促进增殖、分化的黑色素瘤细胞表型,并且对于 SMAD 或 YAP/TAZ 诱导的表型转换是必需的,但不是充分的。这些结果确定了黑色素瘤表型转换背后的信号通路之间的上位相互作用,并强调了黑色素瘤治疗靶点的优先级。