Wadsworth Center, New York State Department of Health, Albany, NY, USA.
Courant Institute and Department of Biology, New York University, New York, NY, USA; Department of Mathematics and the NSF-Simons Center for Multiscale Cell Fate Research, University of California, Irvine, Irvine, CA, USA.
Curr Biol. 2022 Mar 14;32(5):1049-1063.e4. doi: 10.1016/j.cub.2022.01.013. Epub 2022 Feb 1.
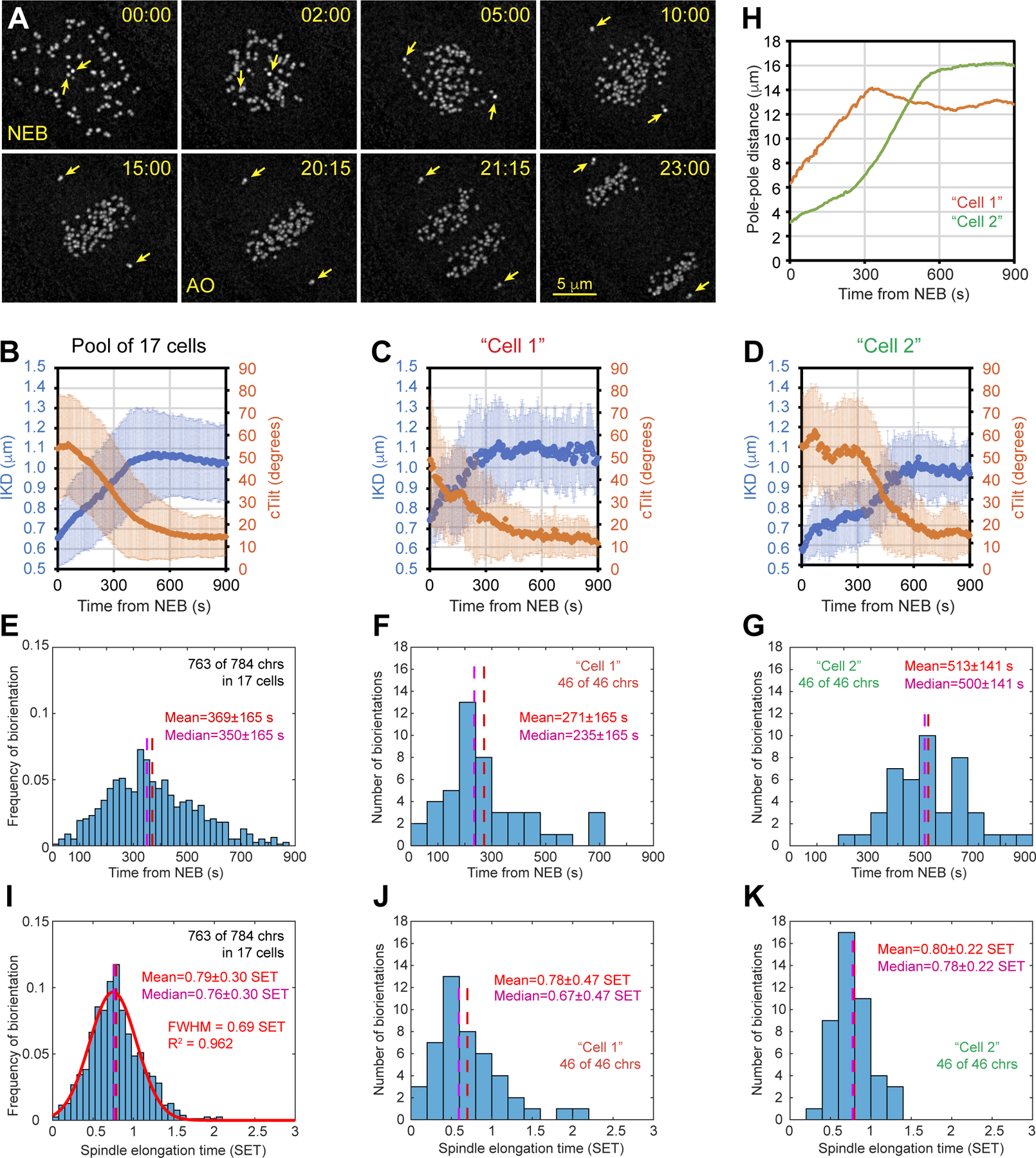
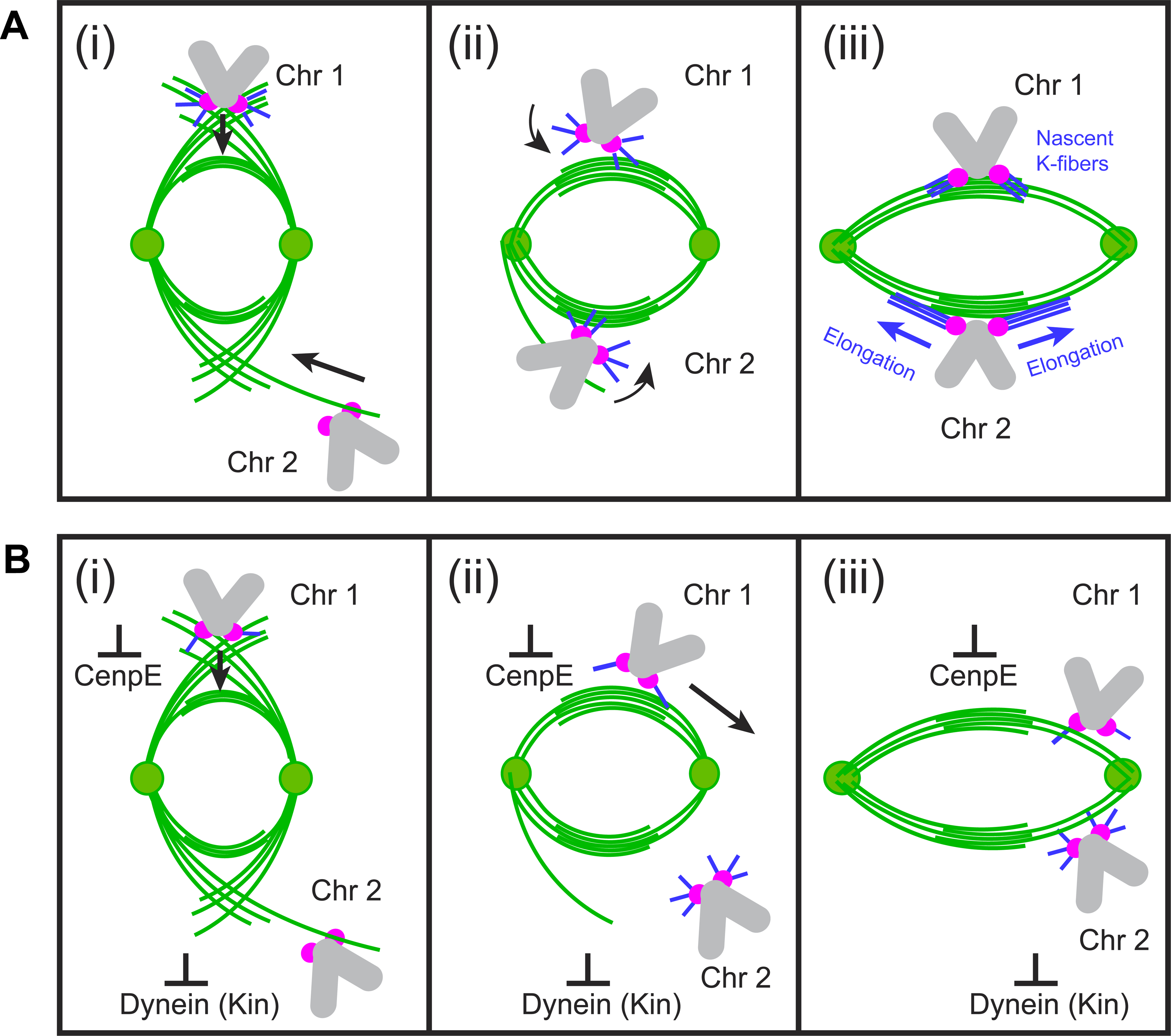
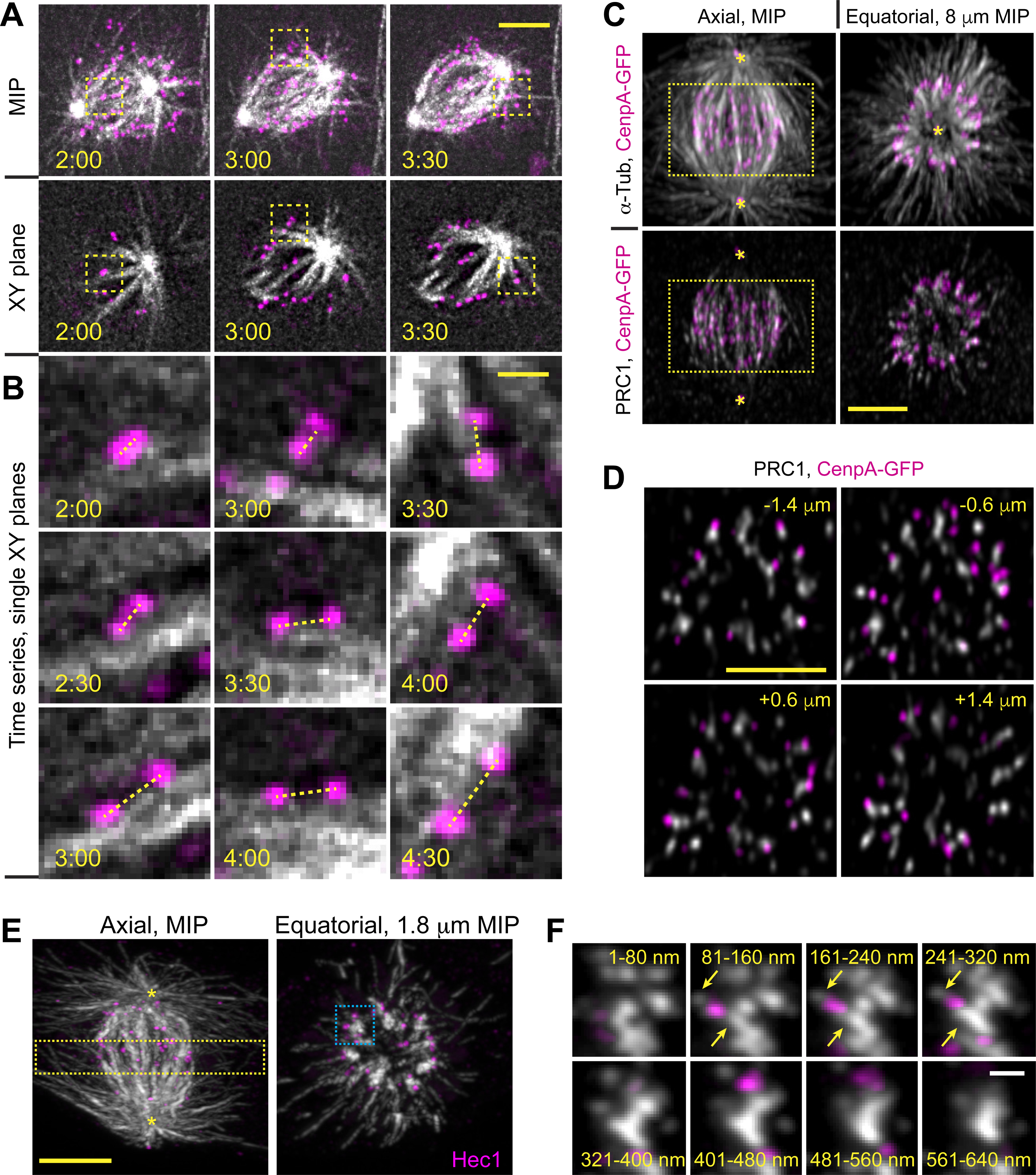
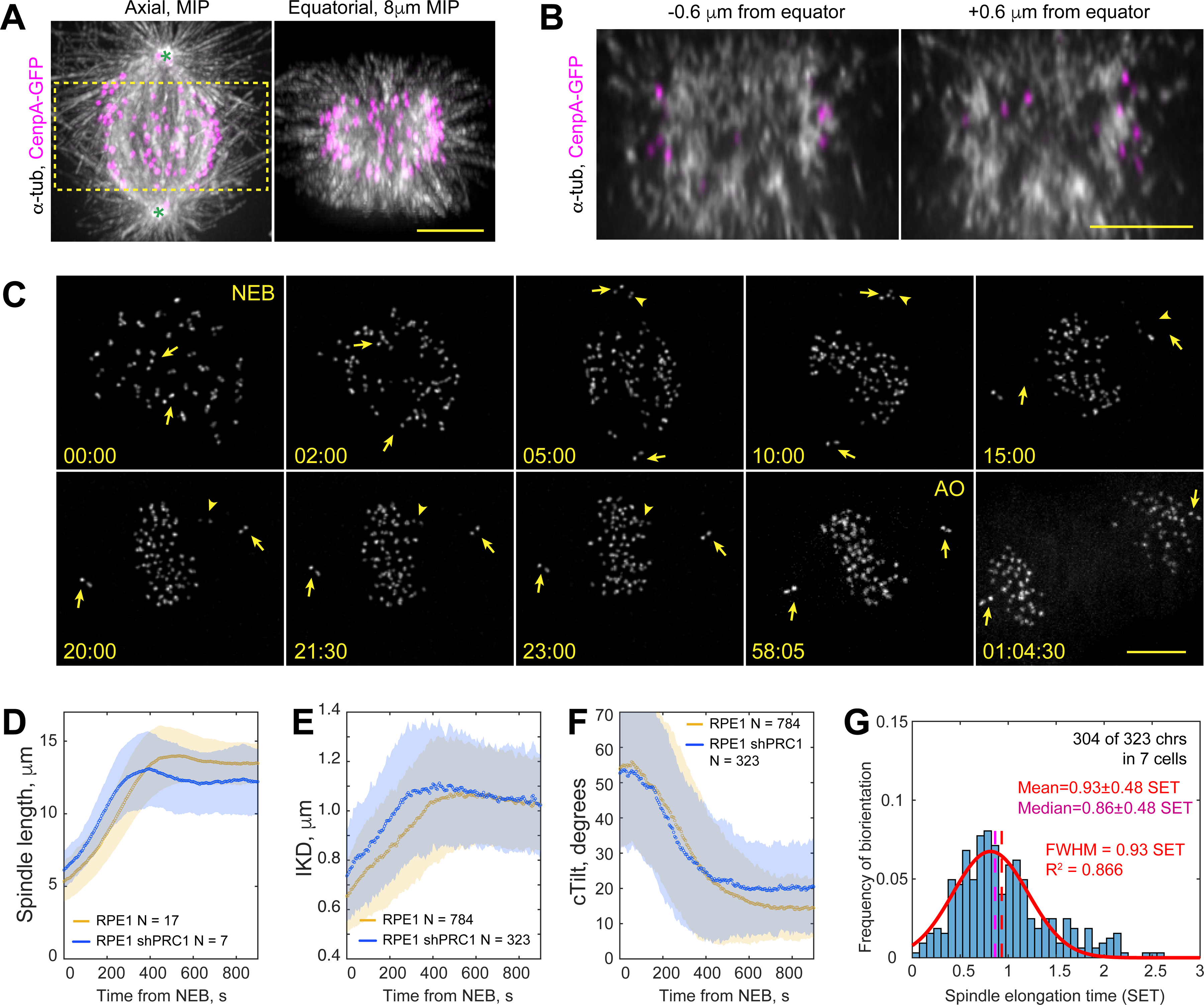
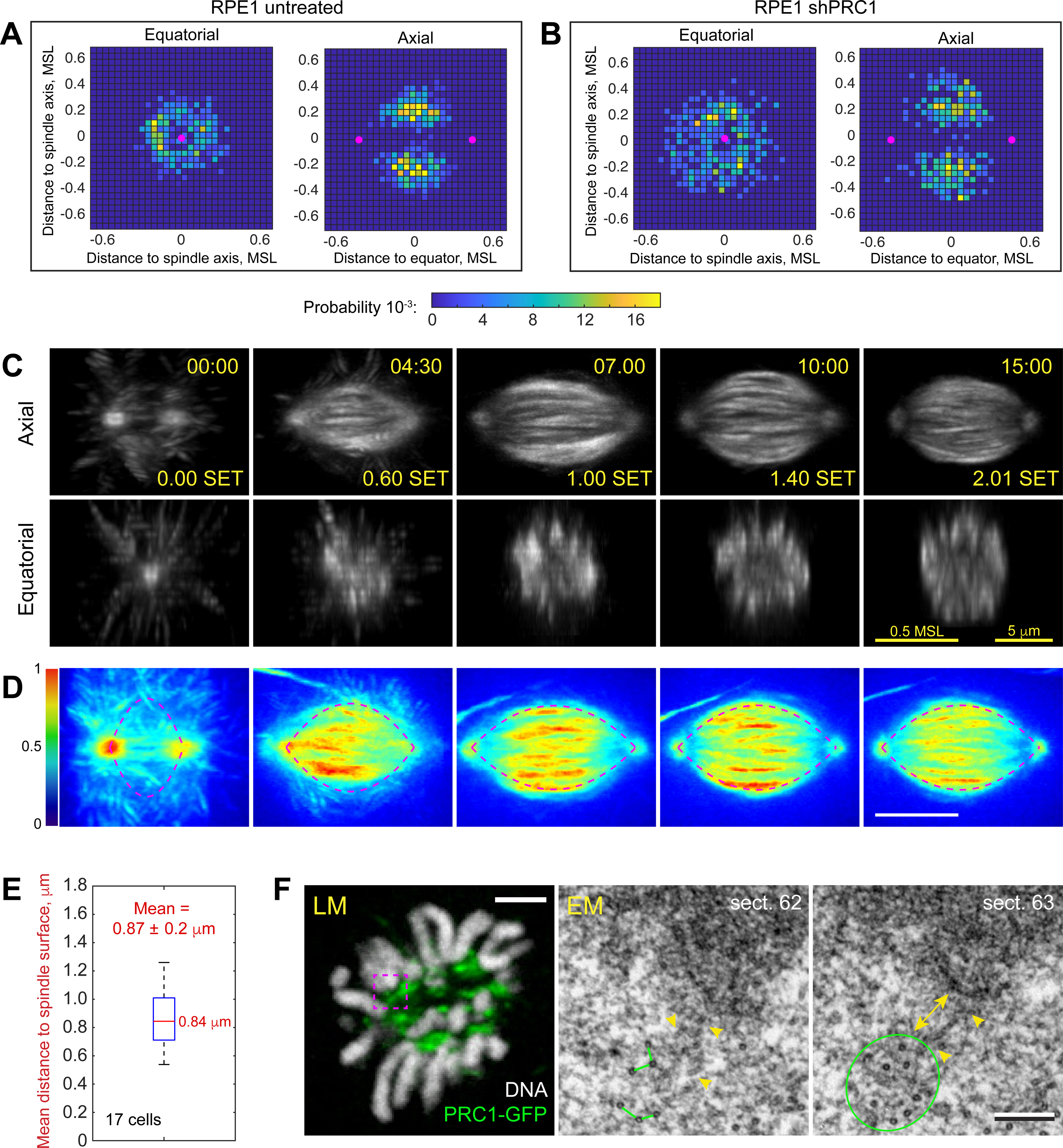
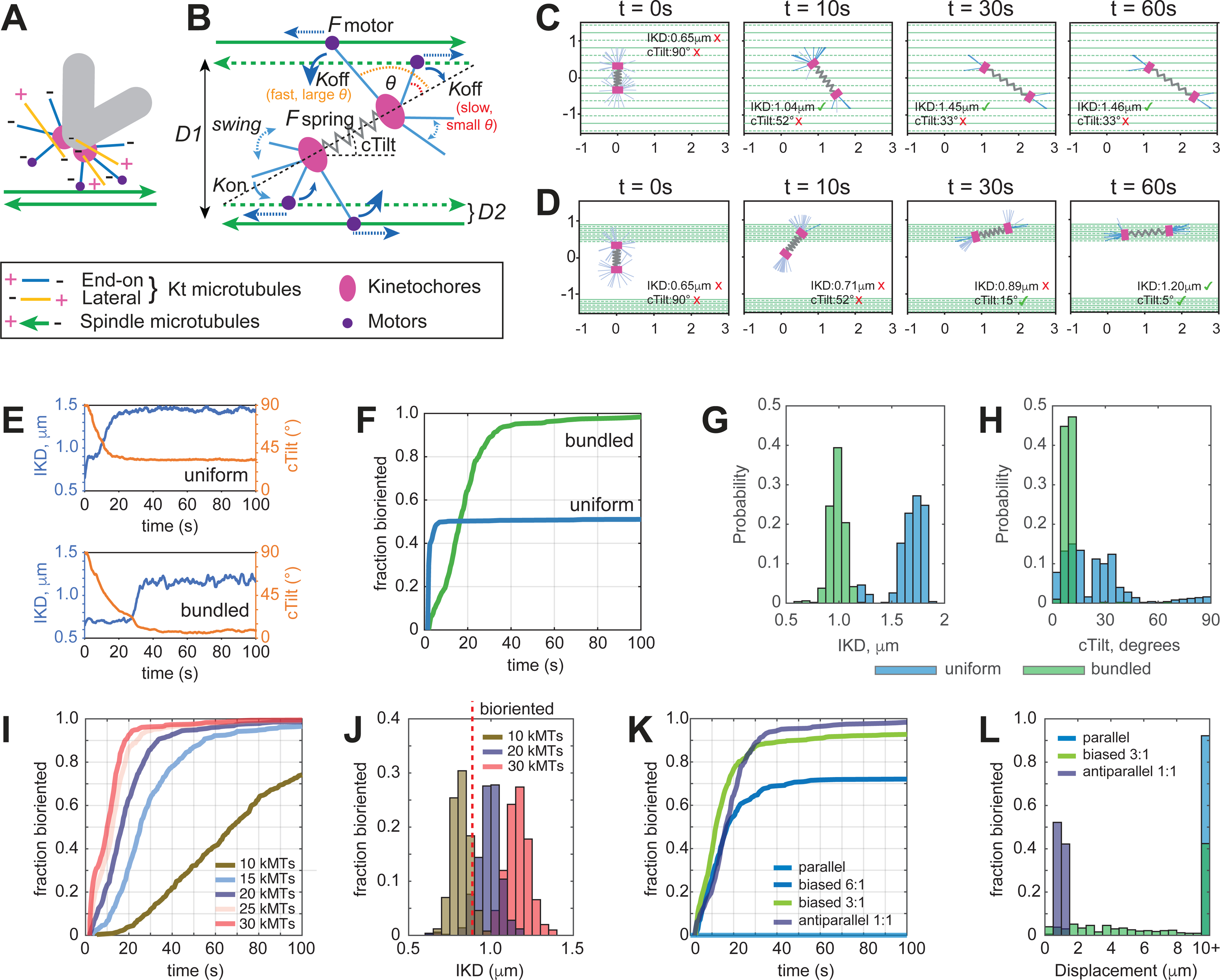
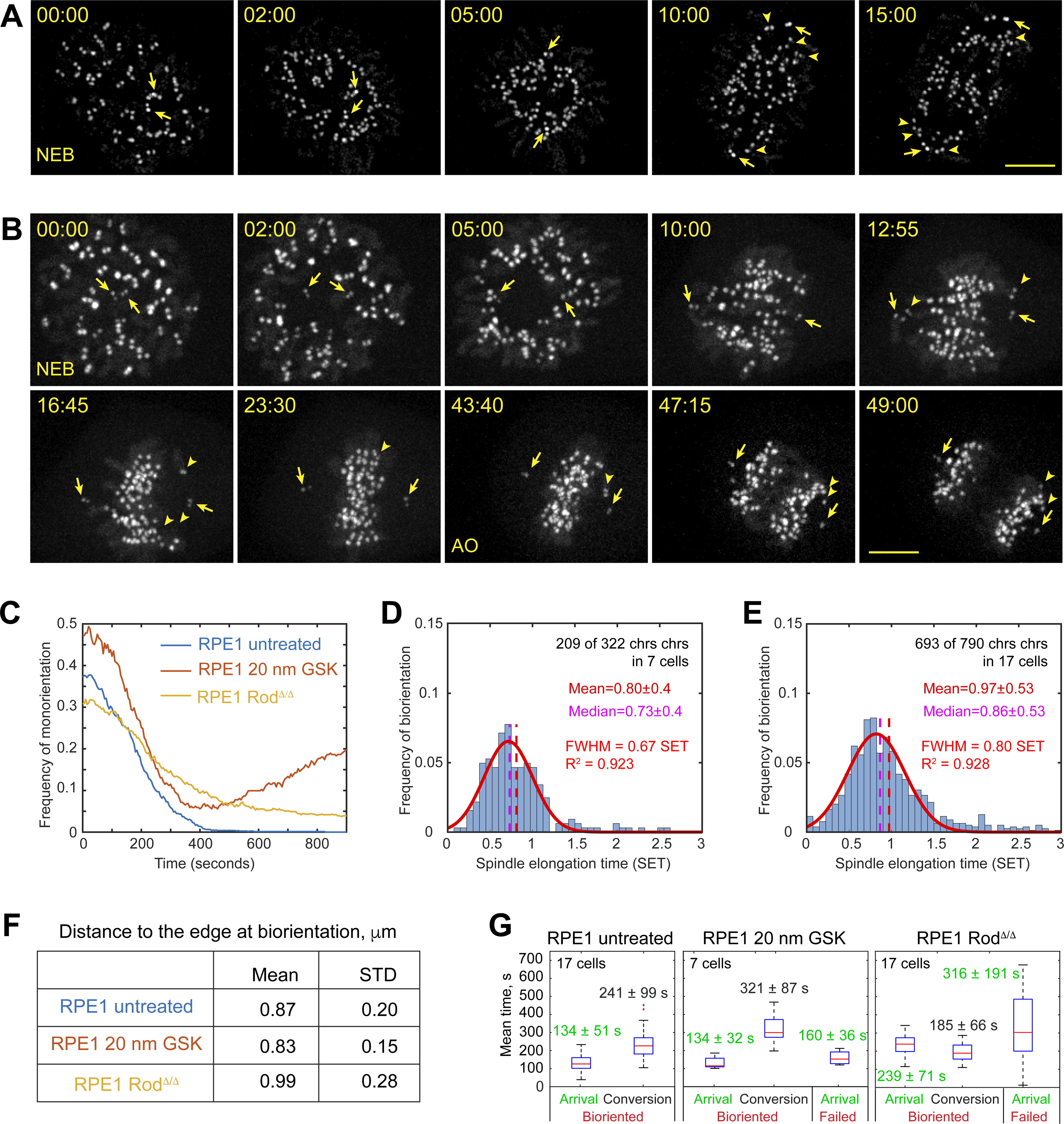
Proper segregation of chromosomes during mitosis depends on "amphitelic attachments"-load-bearing connections of sister kinetochores to the opposite spindle poles via bundles of microtubules, termed as the "K-fibers." Current models of spindle assembly assume that K-fibers arise largely from stochastic capture of microtubules, which occurs at random times and locations and independently at sister kinetochores. We test this assumption by following the movements of all kinetochores in human cells and determine that most amphitelic attachments form synchronously at a specific stage of spindle assembly and within a spatially distinct domain. This biorientation domain is enriched in bundles of antiparallel microtubules, and perturbation of microtubule bundling changes the temporal and spatial dynamics of amphitelic attachment formation. Structural analyses indicate that interactions of kinetochores with microtubule bundles are mediated by non-centrosomal short microtubules that emanate from most kinetochores during early prometaphase. Computational analyses suggest that momentous molecular motor-driven interactions with antiparallel bundles rapidly convert these short microtubules into nascent K-fibers. Thus, load-bearing connections to the opposite spindle poles form simultaneously on sister kinetochores. In contrast to the uncoordinated sequential attachments of sister kinetochores expected in stochastic models of spindle assembly, our model envisions the formation of amphitelic attachments as a deterministic process in which the chromosomes connect with the spindle poles synchronously at a specific stage of spindle assembly and at a defined location determined by the spindle architecture. Experimental analyses of changes in the kinetochore behavior in cells with perturbed activity of molecular motors CenpE and dynein confirm the predictive power of the model.
在有丝分裂过程中,染色体的正确分离依赖于“双联体附着”——通过微管束将姐妹动粒连接到相对纺锤极的承重连接,这些微管束被称为“K 纤维”。目前的纺锤体组装模型假设 K 纤维主要来自微管的随机捕获,这种捕获在随机的时间和位置发生,并且在姐妹动粒上独立发生。我们通过跟踪人类细胞中所有动粒的运动来检验这一假设,并确定大多数双联体附着在纺锤体组装的特定阶段同步形成,并在空间上明显不同的区域形成。这个双取向域富含平行微管束,微管束束的扰动改变了双联体附着形成的时空动力学。结构分析表明,动粒与微管束的相互作用是由非中心体的短微管介导的,这些短微管在早中期从大多数动粒中伸出。计算分析表明,与平行束的重要分子马达驱动相互作用迅速将这些短微管转化为新生的 K 纤维。因此,姐妹动粒的承重连接同时在相对纺锤极上形成。与纺锤体组装的随机模型中预期的姐妹动粒不协调的顺序附着相反,我们的模型将双联体附着的形成视为一个确定性过程,其中染色体在纺锤体组装的特定阶段同步连接到纺锤体极,并在由纺锤体结构决定的特定位置连接。对受分子马达 CenpE 和动力蛋白活性扰动的细胞中动粒行为变化的实验分析证实了该模型的预测能力。